Restablecimiento de los gradientes iónicos de sodio y potasio tras completarse los potenciales de acción: la importancia del metabolismo de la energía
La propagación de cada potencial de acción a lo largo de una fibra nerviosa reduce ligeramente las diferencias de concentración de sodio y de potasio en el interior y en el exterior de la membrana, porque los iones sodio difunden hacia el interior durante la despolarización y los iones potasio difunden hacia el exterior durante la repolarización.
Una característica especial de la bomba Na+-K+-adenosina trifosfatasa es que su grado de actividad se estimula mucho cuando se acumula un exceso de iones sodio en el interior de la membrana celular. De hecho, la actividad de bombeo aumenta aproximadamente en proporción a la tercera potencia de esta concentración intracelular de sodio.
Meseta en algunos potenciales de acción
En algunos casos la membrana excitada no se repolariza inmediatamente después de la despolarización; por el contrario, el potencial permanece en una meseta cerca del máximo del potencial de espiga durante muchos milisegundos, y solo después comienza la repolarización

La causa de la meseta es una combinación de varios factores.
En primer lugar, en el proceso de despolarización del músculo cardíaco participan dos tipos de canales:
- 1) los canales de sodio habituales activados por el voltaje, denominados canales rápidos
- 2) los canales de calcio-sodio activados por el voltaje (canales de calcio de tipo L), que tienen una apertura lenta y que, por tanto, se denominan canales lentos.
. La meseta termina cuando se cierran los canales de calcio-sodio y aumenta la permeabilidad a los iones potasio.
Ritmicidad de algunos tejidos excitables: descarga repetitiva
Las descargas repetitivas autoinducidas aparecen normalmente en el corazón, en la mayor parte del músculo liso y en muchas neuronas del sistema nervioso central. Estas descargas rítmicas producen:
- 1) el latido rítmico del corazón
- 2) el peristaltismo rítmico de los intestinos
- 3) fenómenos neuronales, como el control rítmico de la respiración.

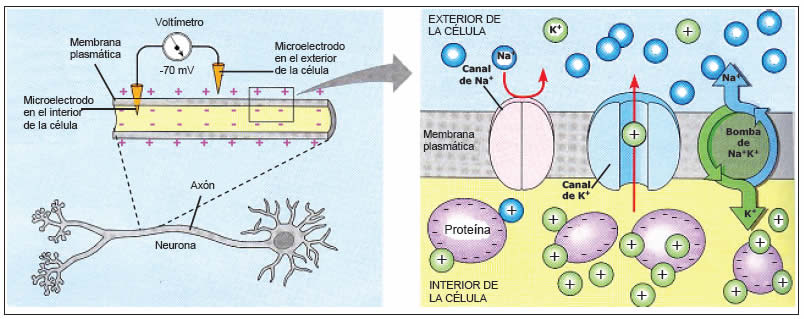
Proceso de reexcitación necesario para la ritmicidad espontánea
Para que se produzca ritmicidad espontánea la membrana, incluso en su estado natural, debe ser lo suficientemente permeable a los iones sodio (o a los iones calcio y sodio a través de los canales lentos de calcio-sodio) como para permitir la despolarización automática de la membrana.
Por tanto, se produce la siguiente secuencia:
- 1) algunos iones sodio y calcio fluyen hacia el interior
- 2) esta actividad produce aumento del voltaje de la membrana en dirección positiva, que aumenta más la permeabilidad de la membrana
- 3) se produce flujo de entrada de aún más iones
- 4) aumenta más la permeabilidad, de manera progresiva, hasta que se genera un potencial de acción. Después, al final del potencial de acción se repolariza la membrana.
Características especiales de la transmisión de señales en los troncos nerviosos
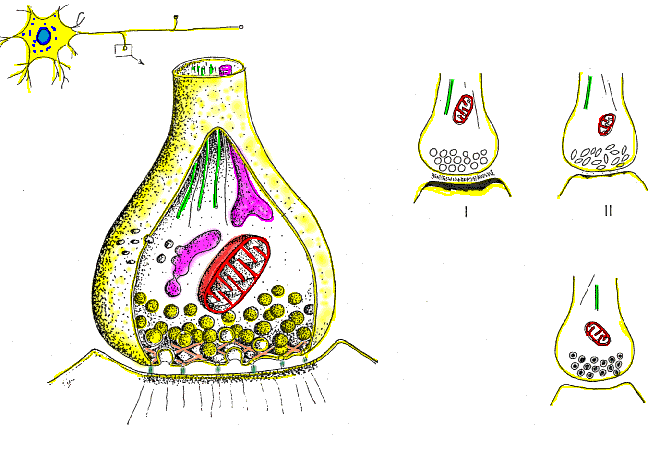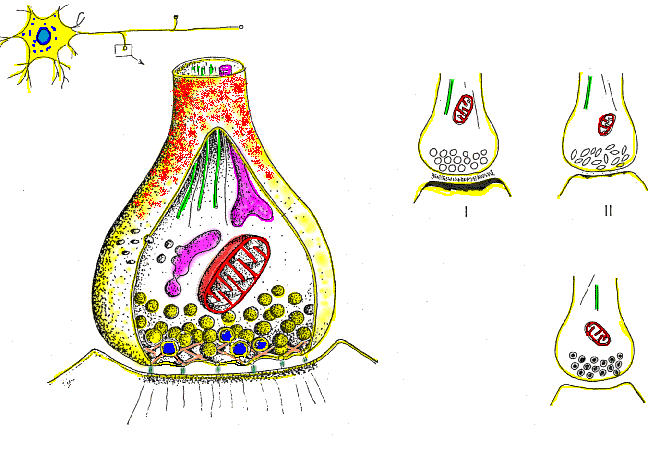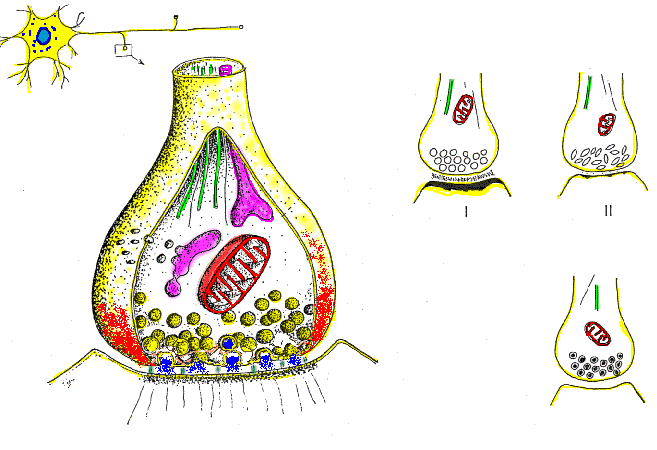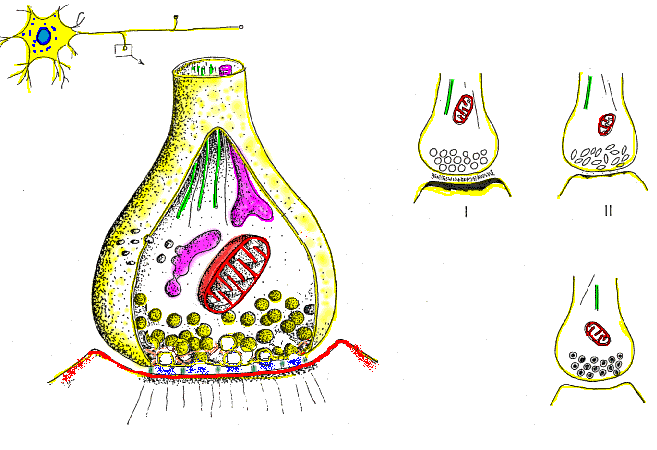
Las fibras grandes son mielinizadas y las pequeñas no mielinizadas. Un tronco nervioso medio contiene aproximadamente el doble de fibras no mielinizadas que mielinizadas.

El núcleo central de la fibra es el axón, y la membrana del axón es la membrana que realmente conduce el potencial de acción. El axón contiene en su centro el axoplasma, que es un líquido intracelular viscoso. Alrededor del axón hay una vaina de
mielina que con frecuencia es mucho más gruesa que el propio axón. Aproximadamente una vez cada 1 a 3 mm a lo largo de la vaina de mielina hay un nódulo de Ranvier.
Las células de Schwann depositan la vaina de mielina alrededor del axón de la siguiente manera: en
primer lugar, la membrana de una célula de Schwann rodea el axón. Después, la célula de Schwann rota muchas veces alrededor del axón, depositando múltiples capas de membrana de la célula de Schwann que contiene la sustancia lipídica esfingomielina. Esta sustancia es un excelente aislante eléctrico que disminuye el flujo iónico a través de la membrana aproximadamente 5.000 veces. En la unión entre dos células de Schwann sucesivas a lo largo del axón permanece una pequeña zona no aislada de solo 2 a 3 μm de longitud en la que los iones pueden seguir fluyendo con facilidad a través de la membrana del axón entre el líquido extracelular y el líquido intracelular del interior del axón. Esta zona se denomina nódulo de Ranvier.

Conducción «saltatoria» en las fibras mielinizadas de un nódulo a otro
La corriente eléctrica fluye por el líquido extracelular circundante que está fuera de la vaina de mielina, así como por el axoplasma del interior del axón, de un nódulo a otro, excitando nódulos sucesivos uno después de otro. Así, el impulso nervioso recorre a saltos la fibra, lo que es el origen del término «saltatoria».

La conducción saltatoria es útil por dos motivos:
- Primero, al hacer que el proceso de despolarización salte intervalos largos a lo largo del eje de la fibra nerviosa, este mecanismo aumenta la velocidad de la transmisión nerviosa en las fibras mielinizadas hasta 5 a 50 veces
- Segundo, la conducción saltatoria conserva la energía para el axón porque solo se despolarizan los nódulos, permitiendo una pérdida de iones tal vez 100 veces menor de lo que sería necesario de otra manera
Por tanto precisa poco gasto de energía para restablecer las diferencias de concentración de sodio y de potasio a través de la membrana después de una serie de impulsos nerviosos.
Velocidad de conducción en las fibras nerviosas:
La velocidad de conducción del potencial de acción en las fibras nerviosas varía desde tan solo 0,25 m/s en las fibras no mielinizadas pequeñas hasta 100 m/s (un valor superior a la longitud de un campo de fútbol en un segundo) en las fibras mielinizadas grandes.
