Funciones motoras de la médula espinal: los reflejos medulares
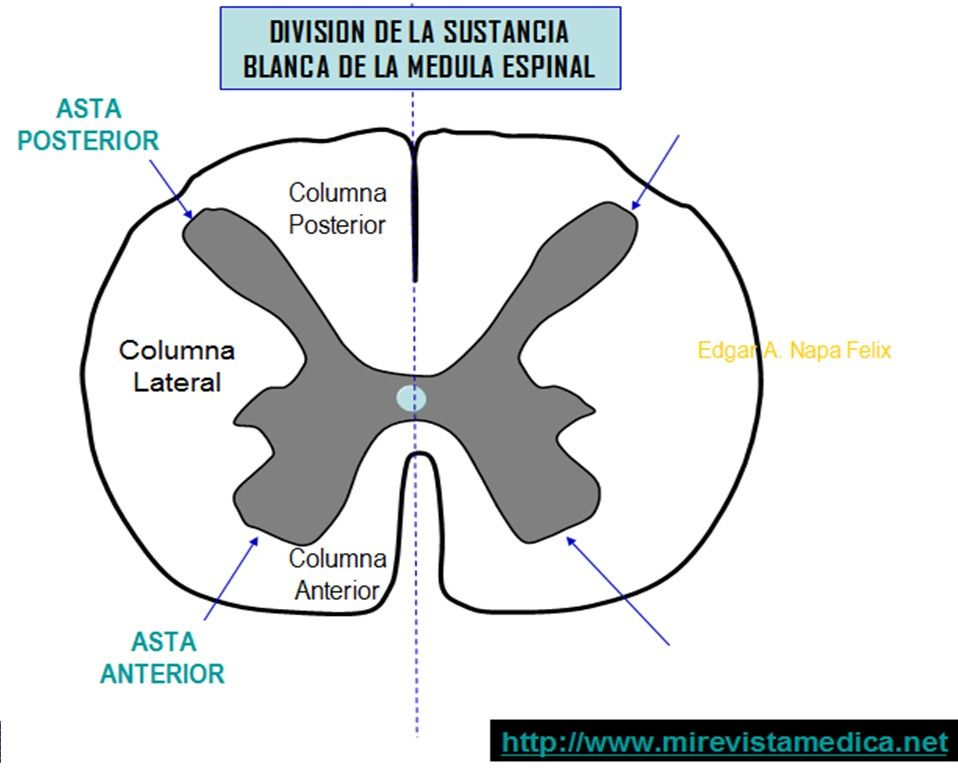
La sustancia gris medular es la zona de integración para los reflejos medulares.
Las señales sensitivas entran por las raíces posteriores o dorsales, posteriormente estas señales viajan hacia dos destinos diferentes:
- A la sustancia gris de la médula suscitando reflejos medulares segmentarios de ámbito local.
- A zonas superiores como el tronco del encéfalo o incluso la corteza cerebral,
Cualquier segmento de la médula espinal contiene:
- Neuronas sensitivas de relevo
- Motoneuronas anteriores
- Interneuronas
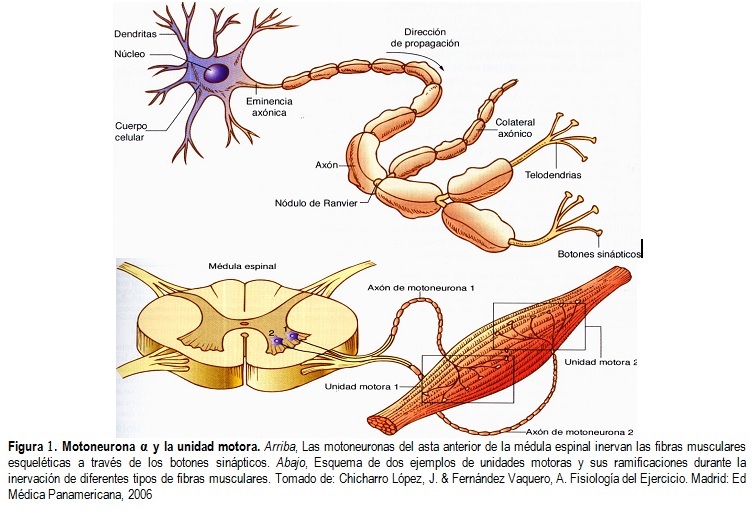
Motoneuronas anteriores
Se encuentran en las astas anteriores de la sustancia gris medular y de ellas nacen las fibras nerviosas que salen de la médula a través de las raíces anteriores e inervan directamente las fibras de los músculos esqueléticos.
son de dos tipos:
- motoneuronas α
- motoneuronas γ.
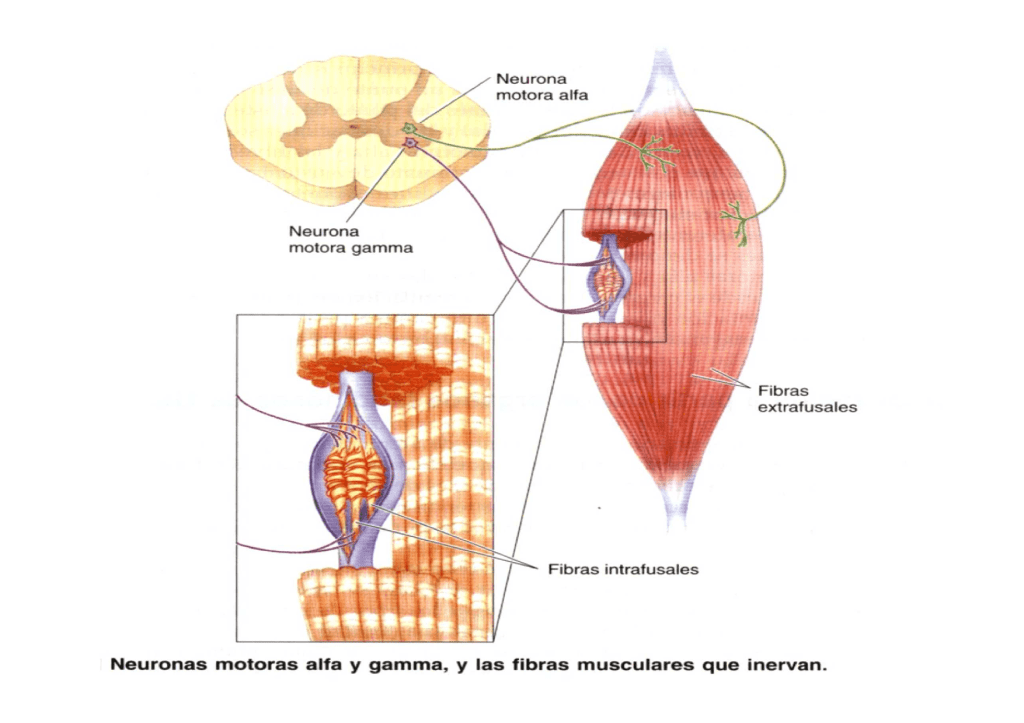
Motoneuronas α
Las motoneuronas α dan origen a unas fibras nerviosas motoras grandes de tipo Aα que al entrar en el músculo inervan las grandes fibras musculares esqueléticas.

Motoneuronas γ
Son mucho más pequeñas y que están situadas en las astas anteriores de la médula espinal, se encargan de transmitir impulsos a través de unas fibras nerviosas motoras γ de tipo A (Aγ) dirigidas hacia las fibras intrafusales que ocupan el centro del huso muscular y se encargan de controlar el «tono» básico del músculo
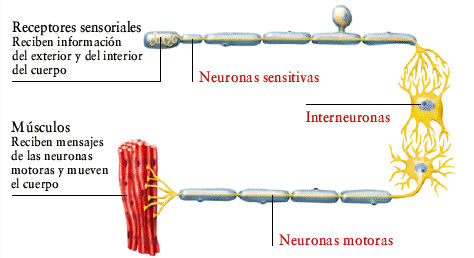
Interneuronas:
Están presentes en todas las regiones de la sustancia gris medular, en las astas posteriores, las astas anteriores y las zonas intermedias, son más numerosas, pequeñas y poseen una naturaleza muy excitable.
Las conexiones entre las interneuronas y las motoneuronas anteriores son las responsables de la mayoría de las funciones integradoras que cumple la médula espinal.
los distintos tipos de circuitos neuronales que existen en los grupos de interneuronas presentes en la médula espinal son:
- Divergentes
- Convergentes
- Descarga repetida
Las células de Renshaw:
Son células inhibidoras que transmiten señales de este carácter hacia las motoneuronas adyacentes, se encuentran en en las astas anteriores de la médula espinal.
Por tanto, la estimulación de cada motoneurona tiende a inhibir a las motoneuronas contiguas según un efecto denominado inhibición lateral; estas permiten la transmisión sin mengua de la señal primaria en la dirección deseada a la vez que se suprime la tendencia a su dispersión lateral.
Fibras propioespinales: Son las fibras nerviosas que ascienden y descienden por la médula espinal, su recorrido va de un segmento medular a otro y suministran una vía para los reflejos multisegmentarios que se encargan de coordinar los movimientos simultáneos de las extremidades anteriores y posteriores.

Funcionamiento muscular exige no solo la excitación del músculo por parte de las motoneuronas anteriores de la médula espinal, sino también una retroalimentación permanente con la información sensitiva que llega a ella procedente de cualquier músculo, para indicar su estado funcional en cada momento mediante dos tipos especiales de receptores sensitivo:
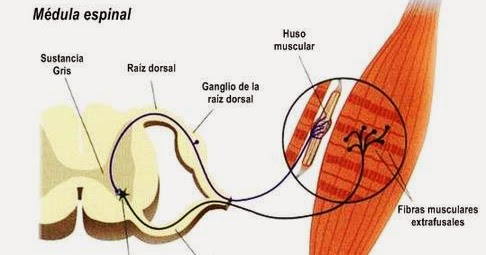
- 1) los husos musculares: se encuentran en el vientre muscular y se encargan de enviar información hacia el sistema nervioso sobre la longitud del músculo o la velocidad con la que varía esta magnitud,
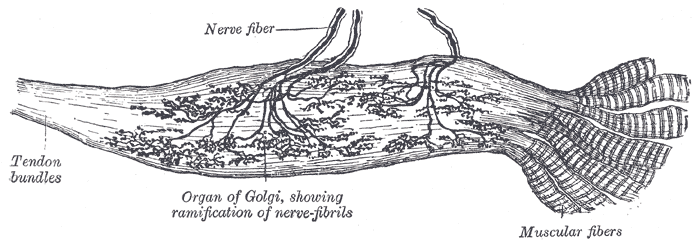
- 2) los órganos tendinosos de Golgi: se encuentran en tendones musculares y transmiten información sobre la tensión tendinosa o su ritmo de cambio.
Estos transmiten una tremenda cantidad de información no solo hacia la médula espinal, sino también hacia el cerebelo e incluso a la corteza cerebral, contribuyendo a que cada una de estas porciones del sistema nervioso intervenga en el control de la contracción muscular.

Función receptora del huso muscular:
Se encuentra dispuesto alrededor de 3 a 12 fibras musculares intrafusales diminutas cuyos extremos acaban en punta y se fijan al glucocáliz de las grandes fibras extrafusales adyacentes correspondientes al músculo esquelético.
Las fibras musculares intrafusales en su área equidistante entre sus dos extremos, contiene pocos filamentos de actina y miosina o ninguno. Por tanto, esta parte central no se contrae pero funciona como un receptor sensitivo.
La porción receptora del huso muscular se localiza en la parte central de una fibra muscular intrafusal y en esta región nacen las fibras sensitivas y su estimulación procede del estiramiento de dicha porción intermedia del huso.
En esta zona receptora central del huso muscular existen dos tipos de terminaciones sensitivas, la terminación aferente primaria y la terminación aferente secundaria.
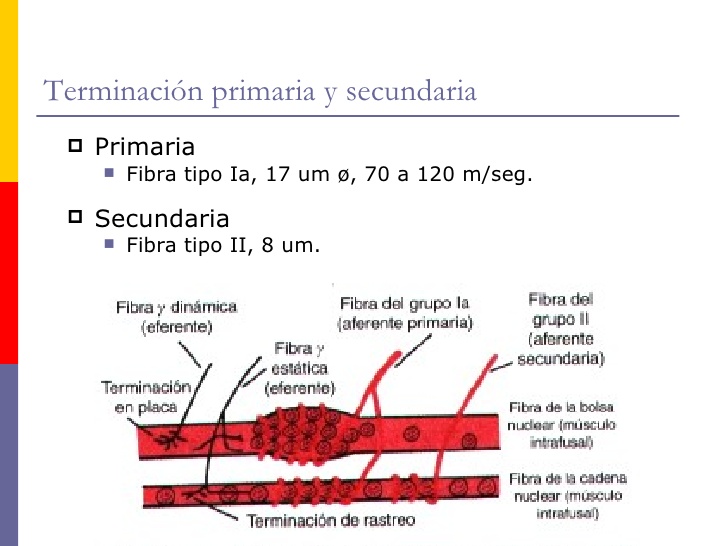
Terminación aferente primaria
Es una gran fibra nerviosa sensitiva rodea la porción central de cada fibra intrafusal se encarga de envía señales sensitivas hacia la médula espinal a una velocidad de 70 a 120 m/s.
Terminación aferente secundaria
Rodea a las fibras intrafusales de la misma forma que lo hace la fibra de tipo Ia
Existen dos tipos de fibras intrafusales en el huso muscular:
- 1) las fibras musculares de bolsa nuclear: varios núcleos de las fibras musculares se encuentran agregados en «bolsas» en la porción central.
- 2) las fibras de cadena nuclear: y cuyos núcleos están alineados formando una cadena a lo largo de toda la región receptora.
Respuesta estática del receptor del huso
Es un efecto que se produce cuando la porción receptora del huso muscular se estira con lentitud y el número de impulsos transmitidos desde las terminaciones primarias y secundarias aumenta casi en proporción directa al grado de estiramiento y las terminaciones continúan transmitiendo estas señales durante varios minutos.
Respuesta dinámica del receptor del huso
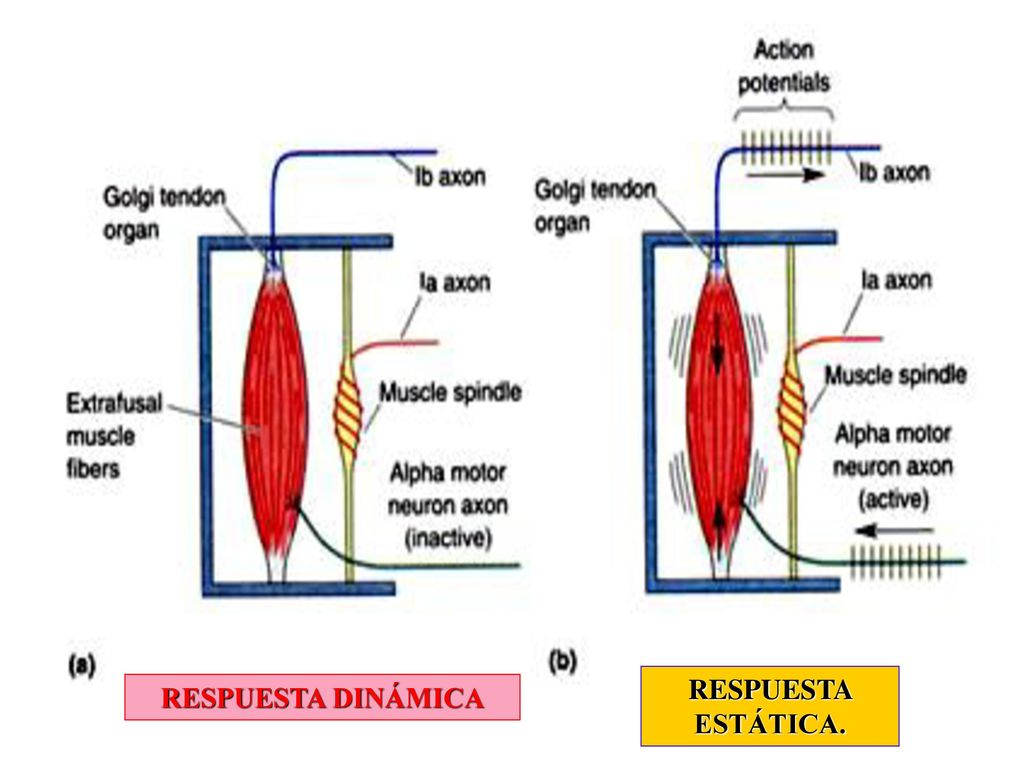
Se produce cuando la longitud del receptor del huso aumenta de forma repentina, la terminación primaria (pero no la secundaria) recibe un estímulo potente, a lo que esta terminación primaria responde de un modo vivísimo a una velocidad de cambio rápida en la longitud del huso.
La terminación primaria manda unos impulsos potentísimos hacia la médula espinal, positivos o negativos, para comunicar cualquier cambio ocurrido en la longitud del receptor del huso.
Existen nervios motores que se dirigen hacia el huso muscular dividiendose en dos tipos:
- γdinámicos (γ-d):excitan sobre todo las fibras intrafusales de bolsa nuclear
- γ-estáticos (γ-s): excitan a lasfibras intrafusales de cadena nuclear
Estos dos tipos de respuestas generados por el huso muscular son importantes en distintas clases de control muscular.
Por ende los husos son capaces de enviar hacia la médula espinal señales positivas (un número mayor de impulsos para indicar el estiramiento muscular) o señales negativas (una cantidad de impulsos reducida para informar

Reflejo miotático muscular
La manifestación más sencilla del funcionamiento del huso es el reflejo miotático o de estiramiento muscular. Siempre que se estira bruscamente un músculo, la activación de los husos causa la contracción refleja de las fibras musculares esqueléticas grandes

El circuito básico del reflejo miotático en el huso muscular se origina por una fibra nerviosa propioceptora de tipo Ia que se origina en un huso muscular y penetra por una raíz posterior de la médula espinal. A continuación, una rama de esta fibra se encamina directamente hacia el asta anterior de la sustancia gris medular y hace sinapsis con las motoneuronas anteriores que devuelven fibras nerviosas motoras al mismo músculo en el que se había originado la fibra del huso citado.
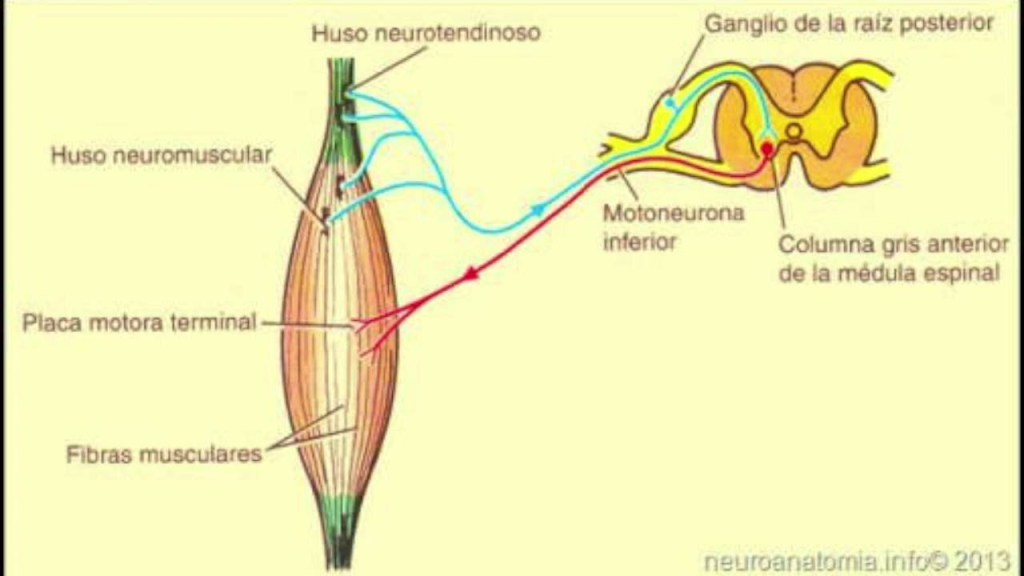
reflejo miotático dinámico
Surge con potentes señales dinámicas transmitidas desde las terminaciones sensitivas primarias de los husos musculares, originada por su estiramiento o distensión rápida
Sirve para oponerse a los cambios súbitos sufridos en la longitud muscular.
reflejo miotático estático
Se mantiene un período prolongado desde que el reflejo miotático dinámico finaliza, produciendo un grado de contracción muscular que puede mantenerse razonablemente constante
Función amortiguadora o suavizadora:Es una función especialmente importante del reflejo miotático ya que evita las oscilaciones o las sacudidas en los movimientos corporales

Coactivación de las motoneuronas α y γ: efecto hace que se contraigan al mismo tiempo las fibras musculares esqueléticas extrafusales y las fibras intrafusales del huso muscular; sus objetivos:
- Evita que varíe la longitud de la porción receptora del huso muscular durante el curso de la contracción muscular completa.
- impide que el reflejo miotático muscular se oponga a la contracción del músculo.
- mantiene la oportuna función amortiguadora del huso
Áreas encefálicas que regulan el sistema motor γ

El sistema eferente γ se activa de forma específica con las señales procedentes de la región facilitadora bulborreticular del tronco del encéfalo y, de un modo secundario, con los impulsos transmitidos hacia la zona bulborreticular desde:
- 1) el cerebelo
- 2) los ganglios basales
- 3) la corteza cerebral.
Una de las funciones más importantes que desempeña el sistema de los husos musculares consiste en estabilizar la posición corporal durante las acciones motoras a tensión.
Esto se realiza debido a la región facilitadora bulborreticular y sus zonas afines del tronco del encéfalo transmiten señales estimuladoras hacia las fibras musculares intrafusales del huso muscular a través de las fibras nerviosas γ, a su vez se acorta los extremos del huso y estira sus regiones receptoras centrales, lo que aumenta la frecuencia de emisión de sus impulsos; obteniendo una articulación sólidamente estabilizada en su posición.
Reflejo tendinoso de Golgi
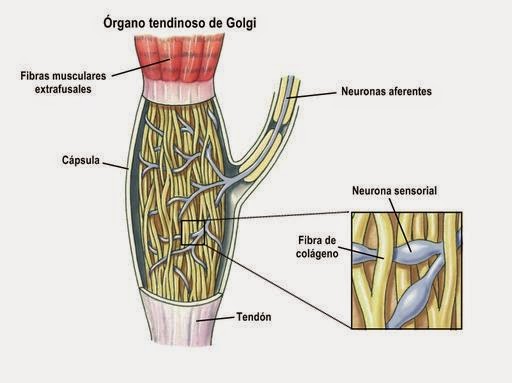
órgano tendinoso de Golgi:
Receptor sensitivo encapsulado por el que pasan las fibras del tendón muscular.
Se estimula cuando este pequeño haz se «tensa» debido a la contracción o el estiramiento del músculo.
Se encarga de identificar la
tensión muscular.
Ofrece una respuesta dinámica y una respuesta estática, siendo potente su reacción cuando la tensión muscular aumenta bruscamente a lo que este receptor aporta al sistema nervioso una información instantánea sobre el grado de tensión en cada pequeño segmento de cualquier músculo.
Transmite su informacion hacia el sistema nervioso central a través de fibras nerviosas grandes de conducción rápida de tipo Ib; enviando sus impulsos hacia las zonas locales de la médula y, después de hacer sinapsis en el asta posterior sigue hacia los fasciculos espinocerebelosos o hacia la corteza cerebral

Ademas de que las señales medulares locales estimulan una sola interneurona inhibidora que actúa sobre la motoneurona anterior. Este circuito local inhibe directamente el músculo correspondiente sin influir sobre los músculos adyacentes.
Las señales que trasmite el organo de Golgi se transmiten hacia la médula espinal para provocar unos efectos reflejos en el músculo correspondiente. Este reflejo tiene un carácter plenamente inhibidor. Por tanto, aporta un mecanismo de retroalimentación negativa que impide la producción de una tensión excesiva en el propio músculo.
Los husos musculares y los órganos tendinosos de Golgi informan a los centros de control motor superiores sobre los cambios instantáneos que tienen lugar en los músculos.
se suscita con mayor potencia mediante la estimulación de las terminaciones para el dolor, como sucede con un pinchazo, el calor o una herida, razón por la que también se le denomina reflejo nociceptivo, o simplemente reflejo al dolor.

Las vías para desencadenar el reflejo flexor alcanzan al conjunto de interneuronas de la médula espinal y solo de un modo secundario las motoneuronas.
La mayoría de las señales de este reflejo atraviesan los siguientes tipos de circuitos básicos:
- 1) circuitos divergentes
- 2) circuitos destinados a inhibir a los músculos antagonistas, llamados circuitos de inhibición recíproca
- 3) circuitos para provocar una posdescarga que dure muchas fracciones de segundo después de finalizar el estímulo.
Además, debido a la posdescarga, el reflejo es capaz de mantener la zona irritada apartada del estímulo durante 0,1 a 3 s después de terminar su acción. Durante este tiempo, otros reflejos y acciones del sistema nervioso
central pueden alejar todo el cuerpo del estímulo doloroso.
Ademas un estímulo doloroso en la cara interna del brazo no solo suscita la contracción de los músculos flexores de esta estructura, sino además la de los abductores para tirar del brazo hacia fuera. Dicho de otro modo, los centros integradores de la médula hacen que se contraigan los músculos que puedan resultar más eficaces para apartar la zona dolorosa del cuerpo del objeto que genera el dolor.
Reacción de apoyo positiva: es tan enérgico que si se pone de pie a un animal cuya médula espinal se haya cortado, a menudo tensa lo suficiente las extremidades como para soportar el peso del cuerpo.
Reacción del imán: . El punto de presión sobre la almohadilla plantar determina la dirección con la que se extenderá el miembro
Reflejo de enderezamiento medular: Cuando un animal espinal está tendido sobre su costado, realizará movimientos descoordinados para tratar de incorporarse

Movimientos de la marcha y la deambulación
Marcha recíproca de las extremidades opuestas: Este efecto deriva de la inervación recíproca existente entre ambos miembros. Cada vez que se den unos pasos en sentido hacia delante con una extremidad, la opuesta corrientemente se desplaza hacia atrás.

Reflejo de «marcar el paso»
En general, los pasos siguen un patrón en diagonal entre las patas delanteras y las traseras. Esta respuesta diagonal constituye otra manifestación de la inervación recíproca, esta vez a lo largo de toda la longitud de la médula hacia arriba y hacia abajo entre las extremidades anteriores y las posteriores
Reflejo de galope:
las extremidades anteriores se desplazan hacia atrás al unísono a la vez que las posteriores se mueven hacia delante. Este reflejo suele suceder cuando se aplican estímulos casi idénticos de estiramiento o de presión a las extremidades de ambos lados del cuerpo al mismo tiempo: su estimulación dispar promueve el reflejo de la marcha en diagonal.
