Músculo cardíaco: el corazón como bomba y la
función de las válvulas cardíacas
Está formado realmente por dos bombas separadas: un corazón derecho que
bombea sangre hacia los pulmones y un corazón izquierdo que bombea sangre a través de la circulación sistémica que aporta flujo sanguíneo a los demás órganos y tejidos del cuerpo.
A su vez, cada uno de estos corazones es una bomba bicameral pulsátil formada por una aurícula y un ventrículo.
Cada una de las aurículas es una bomba débil de cebado del ventrículo, que contribuye a transportar sangre hacia el ventrículo correspondiente.
Los ventrículos después aportan la principal fuerza del bombeo que impulsa la sangre:
- Hacia la circulación pulmonar por el ventrículo derecho,
- Hacia la circulación sistémica por el ventrículo izquierdo.

Mecanismos especiales del corazón producen una sucesión continuada de contracciones cardíacas denominada ritmicidad cardíaca.
Fisiología del músculo cardíaco
El corazón está formado por tres tipos principales de músculo cardíaco: músculo auricular, músculo ventricular y fibras musculares especializadas de excitación y de conducción.
El músculo auricular y ventricular se contrae de manera muy similar al músculo esquelético, excepto porque la duración de la contracción es mucho mayor.
El músculo cardíaco es estriado, igual que el músculo
esquelético.
El músculo cardíaco es estriado, igual que el músculo
esquelético.
Además, el músculo cardíaco tiene las miofibrillas típicas que contienen filamentos de actina y de miosina casi idénticos a los que se encuentran en el músculo esquelético; estos filamentos están unos al lado de otros y se deslizan durante la contracción de la misma manera que ocurre en el músculo esquelético
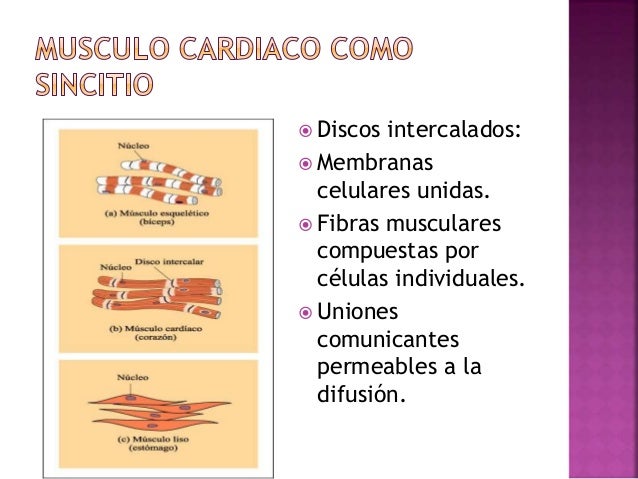
El músculo cardíaco es un sincitio:

Las zonas oscuras que atraviesan las fibras musculares cardíacas se denominan
discos intercalados; realmente son membranas celulares que separan las células musculares cardíacas individuales entre sí.
Es decir, las fibras musculares cardíacas están formadas por muchas células
individuales conectadas entre sí en serie y en paralelo.
En cada uno de los discos intercalados las membranas celulares se fusionan entre sí para formar uniones comunicantes en hendidura permeables que permiten una rápida difusión.
Por tanto, desde un punto de vista funcional los iones se mueven con facilidad en el líquido intracelular a lo largo del eje longitudinal de las fibras musculares cardíacas, de modo que los potenciales de acción viajan fácilmente desde una célula muscular cardíaca a la siguiente, a través de los discos intercalados.
Por tanto, el músculo cardíaco es un sincitio de muchas células musculares cardíacas en el que las células están tan interconectadas entre sí que cuando una célula se excita el potencial de acción se propaga rápidamente a todas.
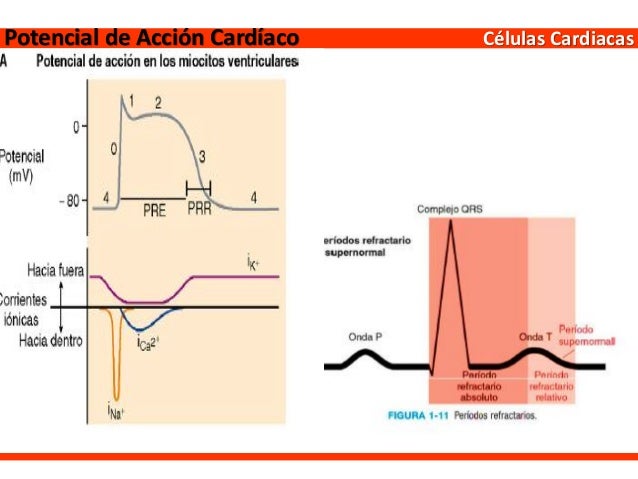
Potenciales de acción en el músculo cardíaco:
El potencial de acción que se registra en una fibra muscular ventricular, es en promedio de aproximadamente 105 mV, lo que significa que el potencial intracelular aumenta desde un valor muy negativo, de aproximadamente –85 mV, entre los latidos hasta un valor ligeramente positivo, de aproximadamente +20 mV, durante cada latido.
Después de la espiga inicial la membrana permanece despolarizada durante aproximadamente 0,2 s, mostrando una
meseta, seguida al final de la meseta de una repolarización súbita.
¿Qué produce el potencial de acción prolongado y la meseta?
Al menos dos diferencias importantes entre las propiedades de la membrana del músculo cardíaco y esquelético son responsables del potencial de acción prolongado y de la meseta del músculo cardíaco.
Primero, el potencial de acción del músculo esquelético está producido casi por completo por la apertura súbita de grandes números de canales rápidos de sodio que permiten que grandes cantidades de iones sodio entren en la fibra muscular esquelética desde el líquido extracelular.
A estos canales se les denomina canales rápidos porque permanecen abiertos solo algunas milésimas de segundo y después se cierran súbitamente.
Al final de este cierre se produce la repolarización y el potencial de acción ha terminado en otra milésima de segundo aproximadamente.
En el músculo cardíaco, el potencial de acción está producido por la apertura de dos tipos de canales:
- Los mismos canales rápidos de sodio activados por el voltaje que en el músculo esquelético
- Otra población totalmente distinta de canales de calcio de tipo L (canales lentos de calcio que también se denominan canales de calcio-sodio.
La segunda diferencia funcional importante entre el músculo cardíaco y el músculo esquelético que ayuda a explicar tanto el potencial de acción prolongado como su meseta es la siguiente: inmediatamente después del inicio del potencial de acción, la permeabilidad de la membrana del músculo cardíaco a los iones potasio disminuye aproximadamente cinco veces, un efecto que no aparece en el músculo esquelético.
Resumen de las fases del potencial de acción del músculo cardíaco
- Fase 0 (despolarización), los canales de sodio rápidos se abren. Cuando la célula cardíaca es estimulada y se despolariza, el potencial de membrana se hace más positivo. Los canales de sodio activados por el voltaje (canales de sodio rápidos) se abren y permiten que el sodio circule rápidamente hacia el interior de la célula y la despolarice. El potencial de membrana alcanza +20 mV aproximadamente antes de que los canales de sodio se cierren.
- Fase 1 (repolarización inicial), los canales de sodio rápidos se cierran. Los canales de sodio se cierran, la célula empieza a repolarizarse y los iones potasio salen de la célula a través de los canale de potasio.
- Fase 2 (meseta), los canales de calcio se abren y los canales de potasio rápidos se cierran. Tiene lugar una breve repolarización inicial y el potencial de acción alcanza una meseta como consecuencia de:1) una mayor permeabilidad a los iones calcio, y 2) la disminución de la permeabilidad a los iones potasio. Los canales de calcio activados por el voltaje se abren lentamente durante las fases 1 y 0, y el calcio entra en la célula. Después, los canales de potasio se cierran, y la combinación de una reducción en la salida de iones potasio y un aumento de la entrada de iones calcio lleva a que el potencial de acción alcance una meseta.
- Fase 3 (repolarización rápida), los canales de calcio se cierran y los canales de potasio lentos se abren. El cierre de los canales iónicos de calcio y el aumento de la permeabilidad a los iones potasio, que permiten que los iones potasio salgan rápidamente de la célula, pone fin a la meseta y devuelve el potencial de membrana de la célula a su nivel de reposo.
- Fase 4 (potencial de membrana de reposo) con valor medio aproximado de −90 mV
Velocidad de la conducción de las señales en el músculo cardíaco:
La velocidad de la conducción de la señal del potencial de acción excitador a lo largo de las fibras musculares auriculares y ventriculares es de unos 0,3 a 0,5 m/s, o aproximadamente 1/250 de la velocidad en las fibras nerviosas grandes y en torno a 1/10 de la velocidad en las fibras musculares esqueléticas.

Período refractario del músculo cardíaco:
El músculo cardíaco, al igual que todos los tejidos excitables, es refractario a la reestimulación durante el potencial de acción.
Por tanto, el período refractario del corazón es el intervalo de tiempo durante el cual un impulso cardíaco normal no puede reexcitar una zona ya excitada de músculo cardíaco. El período refractario normal del ventrículo es de 0,25 a 0,30 s, que es aproximadamente la duración del potencial de acción en meseta prolongado. Hay un período refractario relativo adicional de aproximadamente 0,05 s, durante el cual es más difícil de lo normal excitar el músculo pero, sin embargo, se puede excitar con una señal excitadora muy intensa.
Acoplamiento excitación-contracción: función de los iones calcio y de los túbulos transversos
El término «acoplamiento excitación-contracción» se refiere al mecanismo mediante el cual el potencial de acción hace que las miofibrillas del músculo se contraigan.
Al igual que en el músculo esquelético, cuando un potencial de acción pasa sobre la membrana del músculo cardíaco el potencial de acción se propaga hacia el interior de la fibra muscular cardíaca a lo largo de las membranas de los túbulos transversos (T).
Los potenciales de acción de los túbulos T, a su vez, actúan sobre las membranas de los túbulos sarcoplásmicos longitudinales para producir la liberación de iones calcio hacia el sarcoplasma muscular desde el retículo sarcoplásmico.
Hasta ahora este mecanismo de acoplamiento excitación-contracción es el mismo que el del músculo esquelético, aunque hay un segundo efecto que es bastante diferente.
Además de los iones calcio que se liberan hacia el sarcoplasma desde las cisternas del retículo sarcoplásmico, también difunde una gran cantidad de iones calcio adicionales hacia el sarcoplasma desde los propios túbulos T en el momento del potencial de acción, que abre los canales de calcio dependientes del voltaje a la
membrana del túbulo.
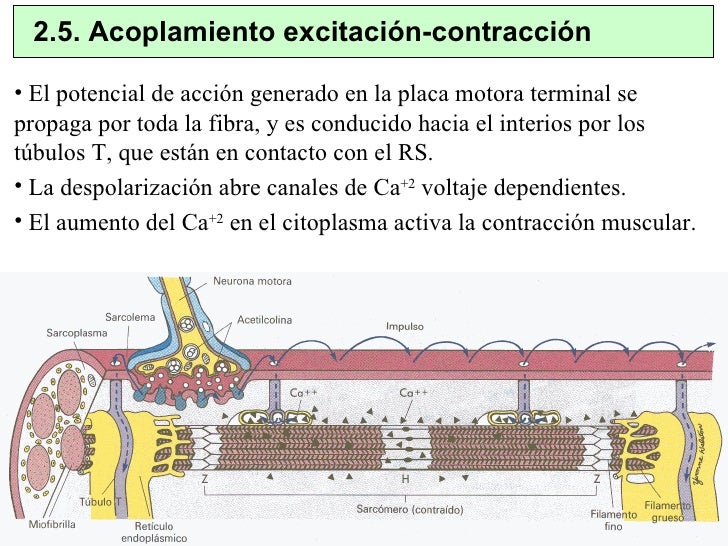
La fuerza de la contracción del músculo cardíaco depende en gran medida de la concentración de iones calcio en los líquidos extracelulares. De hecho, un corazón situado en una solución sin calcio dejará rápidamente de latir.
La razón de esta respuesta es que las aberturas de los túbulos T atraviesan
directamente la membrana de la célula muscular cardíaca hacia los espacios extracelulares que rodean las células, lo que permite que el mismo líquido extracelular que está en el intersticio del músculo cardíaco se introduzca en los túbulos T.
En consecuencia, la cantidad de iones calcio en el sistema de los túbulos T (es decir, la disponibilidad de iones calcio para producir la contracción del
músculo cardíaco) depende en gran medida de la concentración de iones calcio en el líquido extracelular.
En cambio, la fuerza de la contracción del músculo esquelético apenas se ve afectada por cambios moderados de la concentración de calcio en el líquido extracelular porque la contracción del músculo esquelético está producida casi por completo por los iones calcio que son liberados por el retículo sarcoplásmico del interior de la propia fibra muscular esquelética.
Duración de la contracción:
El músculo cardíaco comienza a contraerse algunos milisegundos después de la llegada del potencial de acción y sigue contrayéndose hasta algunos milisegundos después de que finalice. Por tanto, la duración de la contracción del músculo cardíaco depende principalmente de la duración del potencial de acción, incluyendo la meseta, aproximadamente 0,2 s en el músculo auricular y 0,3 s en el músculo ventricular.

Ciclo cardíaco
Los fenómenos cardíacos que se producen desde el comienzo de un latido cardíaco hasta el comienzo del siguiente se denominan ciclo cardíaco.
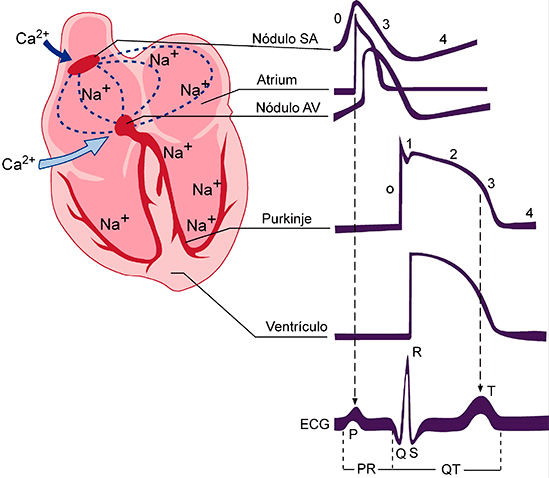
Cada ciclo es iniciado por la generación espontánea de un potencial de acción en el nódulo sinusal.
Este nódulo está localizado en la pared superolateral de la aurícula derecha, cerca del orificio de la vena cava superior, y el potencial de acción viaja desde aquí rápidamente por ambas aurículas y después a través del haz AV hacia los ventrículos.
El ciclo cardíaco está formado por un período de relajación que se denomina diástole, seguido de un período de contracción denominado sístole.
La duración del ciclo cardíaco total, incluidas la sístole y la diástole, es el valor inverso de la frecuencia cardíaca
El aumento de la frecuencia cardíaca reduce la duración del ciclo cardíaco:
El corazón que late a una frecuencia muy rápida no permanece relajado el tiempo suficiente para permitir un llenado completo de las cámaras cardíacas antes de la siguiente contracción.
Relación del electrocardiograma con el ciclo cardíaco:
El electrocardiograma muestra las ondas P, Q, R, S y T
Son los voltajes eléctricos que genera el corazón, y son registrados mediante el
electrocardiógrafo desde la superficie del cuerpo.
- La onda P está producida por la propagación de la despolarización en las aurículas, y es seguid por la contracción auricular, que produce una ligera elevación de la curva de presión auricula inmediatamente después de la onda P electrocardiográfica.
- Por tanto, el complejo QR comienza un poco antes del inicio de la sístole ventricular
- La onda T ventricular representa la fase de repolarización de los ventrículos, cuand las fibras del músculo ventricular comienzan a relajarse. Por tanto, la onda T se produce un poco antes del final de la contracción ventricular.
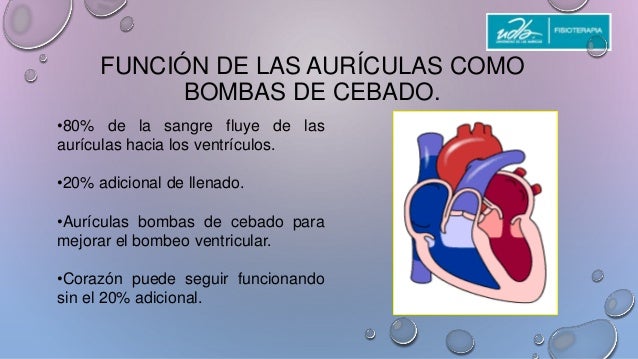
Función de las aurículas como bombas de cebado para los ventrículos
Las aurículas actúan como bombas de cebado que aumentan la eficacia del bombeo ventricular hasta un 20%.
La sangre normalmente fluye de forma continua desde las grandes ventas hacia las aurículas; aproximadamente el 80% de la sangre fluye directamente a través de las aurículas hacia los ventrículos incluso antes de que se contraigan las aurículas.
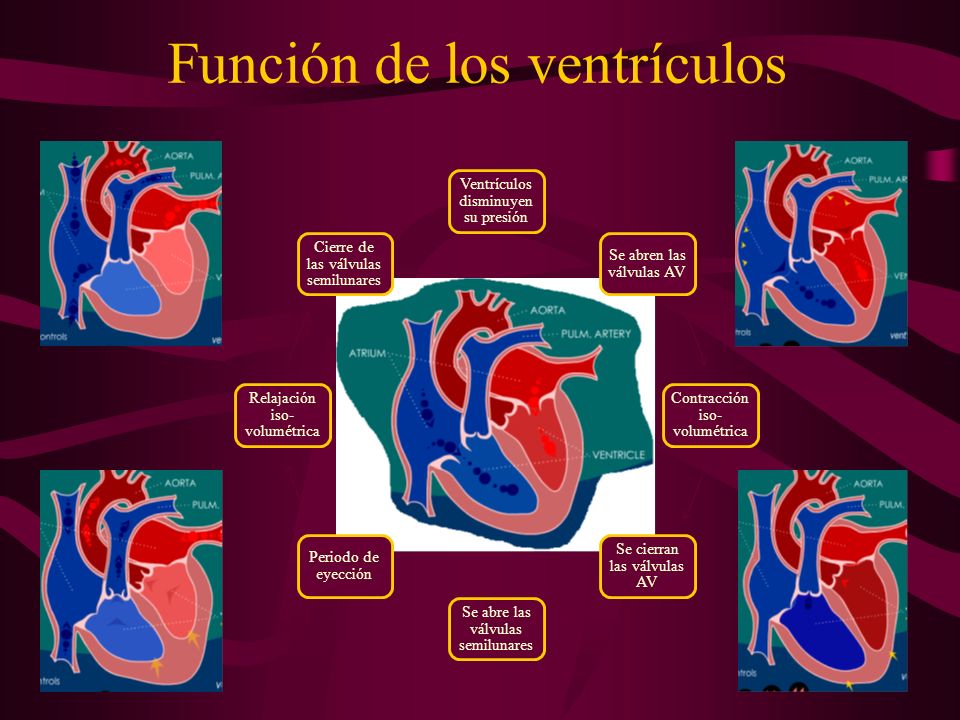
Función de los ventrículos como bombas.:

Durante la sístole ventricular se acumulan grandes cantidades de sangre en las aurículas derecha e izquierda porque las válvulas AV están cerradas.
El período de llenado rápido dura aproximadamente el primer tercio de la diástole. Durante el tercio medio de la diástole normalmente solo fluye una pequeña cantidad de sangre hacia los ventrículos; esta es la sangre que continúa drenando hacia las aurículas desde las venas y que pasa a través de las aurículas directamente hacia los ventrículos.
Desbordamiento de los ventrículos durante la sístole
Período de contracción isovolumétrica (isométrica)
Se produce aumento de la tensión en el músculo cardíaco, pero con un acortamiento escaso o nulo de las fibras musculares.
Inmediatamente después del comienzo de la contracción ventricular se produce un aumento súbito de presión ventricular
Lo que hace que se cierren las válvulas AV.
Después son necesarios otros 0,02 a 0,03 s para que el ventrículo acumule una presión suficiente para abrir las válvulas AV semilunares (aórtica y pulmonar) contra las presiones de la aorta y de la arteria pulmonar.
Por tanto, durante este período se produce contracción en los ventrículos, pero no se
produce vaciado
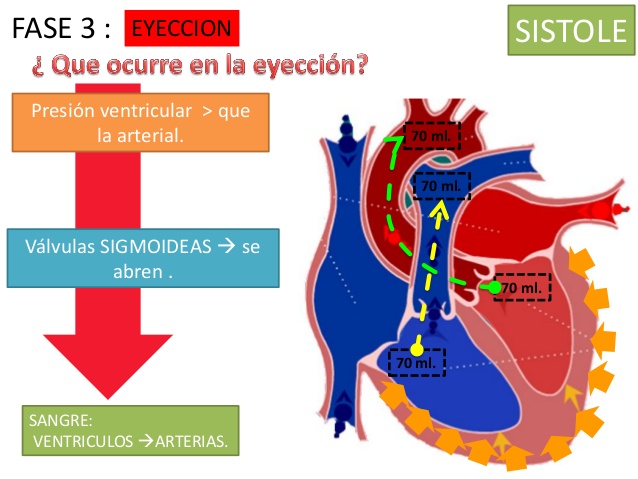
Período de eyección
Cuando la presión ventricular izquierda aumenta ligeramente por encima de 80 mmHg (y la presión ventricular derecha ligeramente por encima de 8 mmHg), las presiones ventriculares abren las válvulas semilunares.
Por tanto, el primer tercio se denomina período de eyección rápida y los dos tercios finales período de eyección lenta.
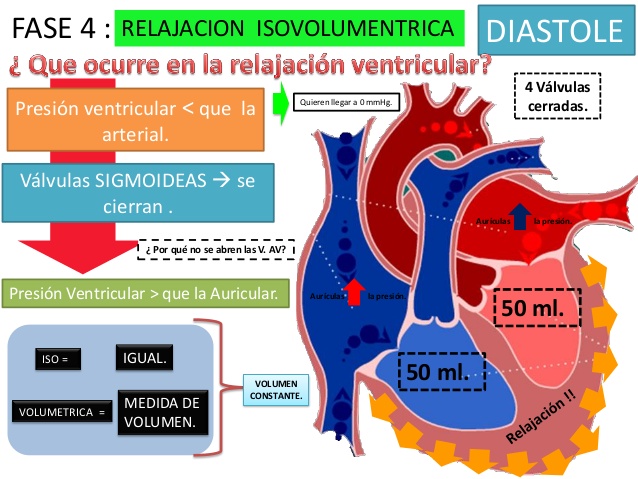
Período de relajación isovolumétrica (isométrica)
Al final de la sístole comienza súbitamente la relajación ventricular, lo que permite que las presiones
intraventriculares derecha e izquierda disminuyan rápidamente.
Las presiones elevadas de las grandes arterias distendidas que se acaban de llenar con la sangre que procede de los ventrículos que se han contraído empujan inmediatamente la sangre de nuevo hacia los ventrículos, lo que cierra súbitamente las válvulas aórtica y pulmonar.
Volumen telediastólico, volumen telesistólico y volumen sistólico:
Durante la diástole, el llenado normal de los ventrículos aumenta el volumen de cada uno de los ventrículos hasta aproximadamente 110 a 120 ml.
Después, a medida que los ventrículos se vacían durante la sístole, el volumen disminuye aproximadamente 70 ml, lo que se denomina volumen sistólico.
El volumen restante que queda en cada uno de los ventrículos, aproximadamente 40 a 50 ml, se denomina volumen telesistólico

Las válvulas cardíacas evitan el flujo inverso de la sangre durante la sístole
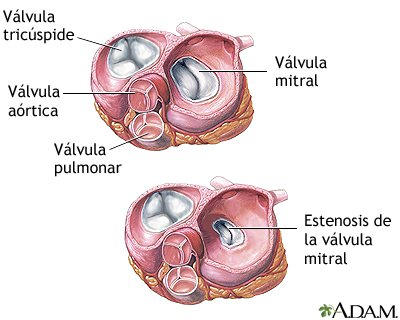
Válvulas auriculoventriculares
Las válvulas AV (las válvulas tricúspide y mitral) impiden el flujo retrógrado de sangre desde los ventrículos hacia las aurículas durante la sístole, y las válvulas semilunares (es decir, las válvulas
aórtica y de la arteria pulmonar) impiden el flujo retrógrado desde las arterias aorta y pulmonar hacia los ventrículos durante la diástole.
Las válvulas AV, que están formadas por una película delgada, casi no precisan ningún flujo retrógrado para cerrarse, mientras que las válvulas semilunares, que son mucho más fuertes, precisan un flujo retrógrado bastante rápido durante algunos milisegundos.
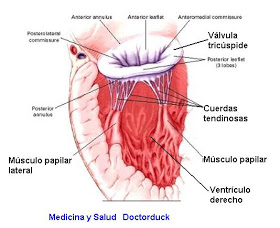
Función de los músculos papilares:
Los músculos papilares se contraen cuando se contraen las paredes ventriculares, pero, al contrario de lo que se podría esperar, no contribuyen al cierre de las válvulas.
Por el contrario, tiran de los velos de las válvulas hacia dentro, hacia los ventrículos, para impedir
que protruyan demasiado hacia las aurículas durante la contracción ventricular
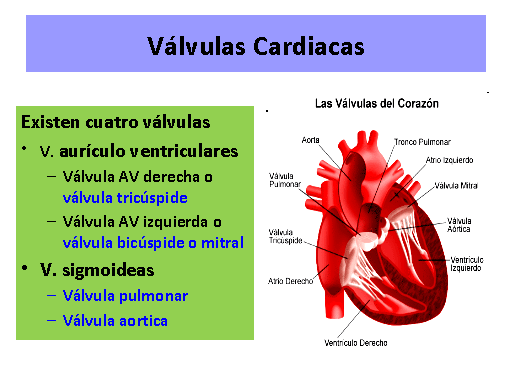
Válvulas aórtica y de la arteria pulmonar.
Debido a sus orificios más pequeños, la velocidad de la eyección de la sangre a través de las válvulas aórtica y pulmonar es mucho mayor que a través de las válvulas AV, que son mucho mayores. Además, debido al cierre rápido y a la eyección rápida, los bordes de las válvulas aórtica y pulmonar están sometidos a una abrasión mecánica mucho mayor que las válvulas AV.
Las válvulas AV tienen el soporte de las cuerdas tendinosas, lo que no ocurre en el caso de las válvulas semilunares.
Curva de presión aórtica
Cuando el ventrículo izquierdo se contrae, la presión ventricular aumenta rápidamente hasta que se abre la válvula aórtica.
Posteriormente, después de que se haya abierto la válvula, la presión del
ventrículo aumenta mucho menos rápidamente; la entrada de sangre en las arterias durante la sístole hace que sus paredes se distiendan y que la
presión aumente hasta aproximadamente 120 mmHg.
Al final de la sístole, después de que el ventrículo izquierdo haya dejado de impulsar sangre y se haya cerrado la válvula aórtica, las paredes elásticas de las arterias mantienen una presión elevada en las arterias, incluso durante la diástole.
Después de que se haya cerrado la válvula aórtica, la presión en el interior de la aorta disminuye lentamente durante toda la sístole porque la sangre que está almacenada en las arterias elásticas distendidas fluye continuamente a través de los vasos periféricos de nuevo hacia las venas.
Análisis gráfico del bombeo ventricular:
- La curva de presión diastólica se determina llenando el corazón con volúmenes de sangre progresivamente mayores y midiendo la presión diastólica inmediatamente antes de que se produzca la contracción ventricular, que es la presión telediastólica del ventrículo.
- La curva de presión sistólica se determina registrando la presión sistólica que se alcanza durante la contracción ventricular a cada volumen de llenado.
- Durante la contracción ventricular, la presión «sistólica» aumenta incluso a volúmenes ventriculares bajos y alcanza un máximo a un volumen ventricular de 150 a 170 ml. Después, a medida que sigue aumentando el volumen, la presión sistólica llega a disminuir en algunas situaciones, como se muestra por la disminución de la curva de presión sistólica
Diagrama volumen-presión durante el ciclo cardíaco: trabajo cardíaco
- Fase I: período de llenado. La fase I del diagrama volumen-presión comienza a un volumen ventricular de aproximadamente 50 ml y una presión diastólica próxima a 2 a 3 mmHg.
- Fase II: período de contracción isovolumétrica. Durante la contracción isovolumétrica el volumen del ventrículo no se modifica porque todas las válvulas están cerradas.
- Fase III: período de eyección. Durante la eyección la presión sistólica aumenta incluso más debido a una contracción aún más intensa del ventrículo. Al mismo tiempo, el volumen del ventrículo disminuye porque la válvula aórtica ya se ha abierto y la sangre sale del ventrículo hacia la aorta.
- Fase IV: período de relajación isovolumétrica. Al final del período de eyección se cierra la válvula aórtica, y la presión ventricular disminuye de nuevo hasta el nivel de la presión diastólica. Así, el ventrículo recupera su valor inicial, en el que quedan aproximadamente 50 ml de sangre en el ventrículo y la presión auricular es de aproximadamente de 2 a 3 mmHg.
Conceptos de precarga y poscarga
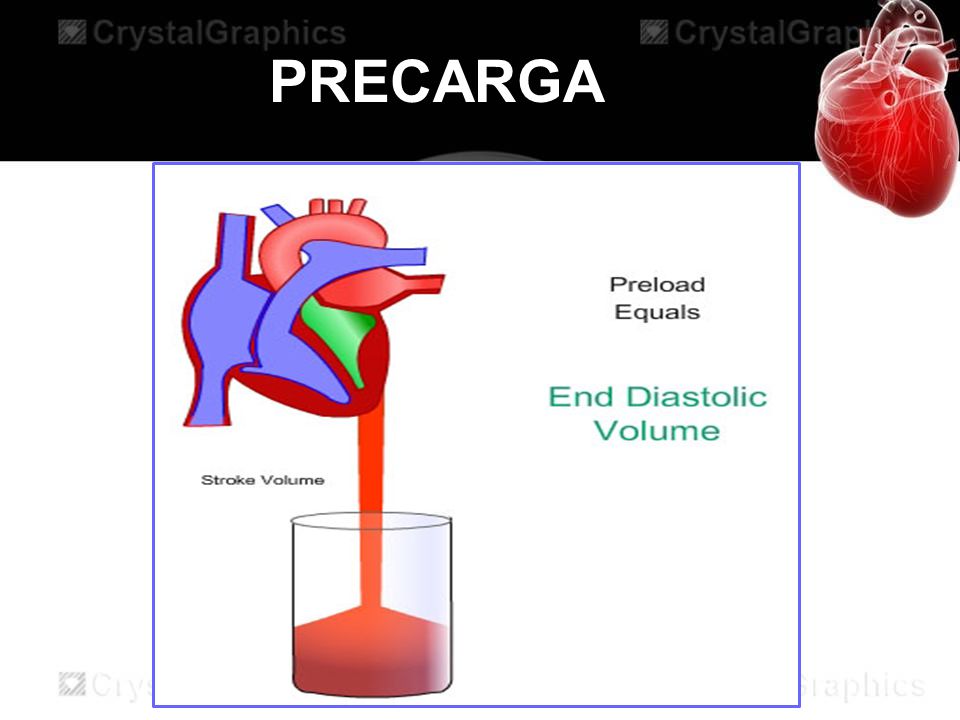
Cuando se evalúan las propiedades contráctiles del músculo es importante especificar el grado de tensión del músculo cuando comienza a contraerse

La poscarga del ventrículo es la presión de la aorta que sale del ventrículo.
(A veces se considera de manera aproximada que la poscarga es la resistencia de la
circulación, en lugar de su presión.)
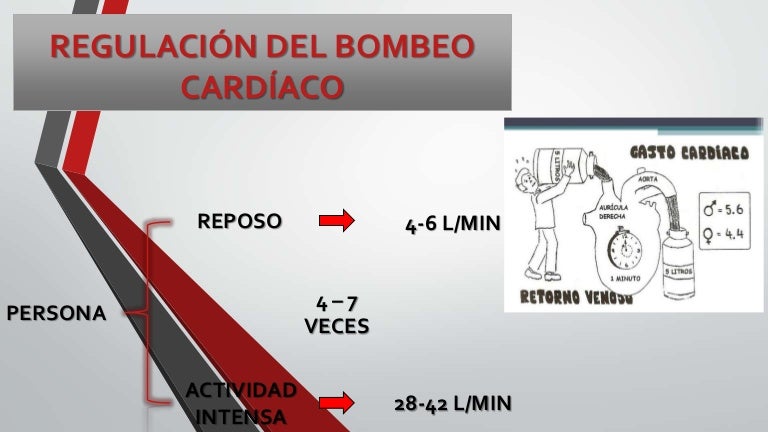
Regulación del bombeo cardíaco
Los mecanismos básicos mediante los que se regula el volumen que bombea el corazón son:
- Regulación cardíaca intrínseca del bombeo en respuesta a los cambios del volumen de la sangre que fluye hacia el corazón
- Control de la frecuencia cardíaca y del bombeo cardíaco por el sistema
- nervioso autónomo
Todos los tejidos periféricos del cuerpo controlan su propio flujo sanguíneo local, y todos los flujos tisulares locales se combinan y regresan a través de las venas hacia la aurícula derecha. El corazón, a su vez, bombea automáticamente hacia las arterias esta sangre que le llega, de modo que pueda fluir de
nuevo por el circuito..
Cuanto más se distiende el músculo cardíaco durante el llenado, mayor es la fuerza de contracción y mayor es la cantidad de sangre que bombea hacia la aorta
¿Cuál es la explicación del mecanismo de Frank-Starling?

Cuando una cantidad adicional de sangre fluye hacia los ventrículos, el propio músculo cardíaco es distendido hasta una mayor longitud. Esta distensión, a su vez, hace que el músculo se contraiga con más fuerza porque los filamentos de actina y de miosina son desplazados hacia un grado más óptimo de superposición para la generación de fuerza. Por tanto, el ventrículo, debido al aumento de la
función de bomba, bombea automáticamente la sangre adicional hacia las arterias.
Una de las mejores formas de expresar la capacidad funcional de los ventrículos de bombear sangre es mediante la curvas de función ventricular.
Medida que los ventrículos se llenan en respuesta a unas presiones auriculares
más altas, se produce aumento del volumen de los dos ventrículos y de la fuerza de la contracción del músculo cardíaco, lo que hace que el corazón bombee mayores cantidades de sangre hacia las arterias.

Control del corazón por los nervios simpáticos y parasimpáticos
La eficacia de la función de bomba del corazón también está controlada por los nervios simpáticos y parasimpáticos (vagos), que inervan de forma abundante el corazón

Mecanismos de excitación del corazón por los nervios simpáticos
La estimulación simpática intensa puede aumentar la frecuencia cardíaca en seres humanos adultos jóvenes desde la frecuencia normal de 70 latidos/min hasta 180 a 200 y, raras veces, incluso 250 latidos/min.
Además, la estimulación simpática aumenta la fuerza de la contracción cardíaca
hasta el doble de lo normal, aumentando de esta manera el volumen de sangre que se bombea y aumentando la presión de eyección. Así, con frecuencia la estimulación simpática puede aumentar el gasto cardíaco máximo hasta dos o tres veces, además del aumento del gasto que produce el mecanismo de Frank-Starling que ya se ha comentado.
Por el contrario, la inhibición de los nervios simpáticos del corazón puede disminuir la función de bomba del corazón en un grado moderado: en condiciones normales, las fibras nerviosas simpáticas que llegan al corazón descargan continuamente a una frecuencia baja que mantiene el bombeo aproximadamente un 30% por encima del que habría sin estimulación simpática.
La estimulación parasimpática (vagal) reduce la frecuencia cardíaca y la fuerza de la contracción

La estimulación intensa de las fibras nerviosas parasimpáticas de los nervios vagos que llegan al corazón puede interrumpir el latido cardíaco durante algunos segundos, pero después el corazón habitualmente «escapa» y late a una frecuencia de 20 a 40 latidos/min mientras continúe la estimulación parasimpática.
Las fibras vagales se distribuyen principalmente por las aurículas y no mucho en los ventrículos, en los que se produce la contracción de potencia del corazón.
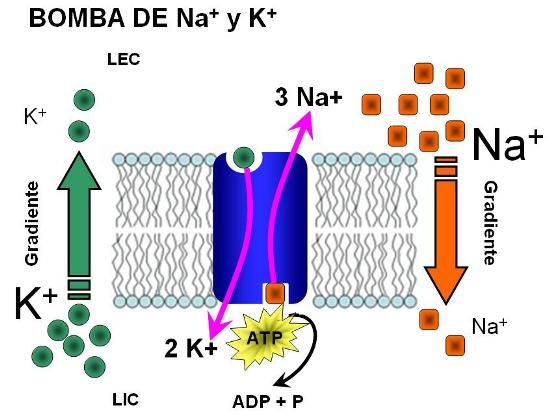
Efecto de los iones potasio y calcio sobre la función cardíaca
Los iones potasio tienen un efecto marcado sobre los potenciales de membrana y desempeñan una función especialmente importante en la activación del proceso contráctil del músculo.
La concentración de cada uno de estos dos iones en los líquidos extracelulares también tenga efectos destacados sobre la función de bomba del corazón

El exceso de potasio hace que el corazón esté dilatado y flácido, y también reduce la frecuencia
cardíaca.
Grandes cantidades de potasio también pueden bloquear la conducción del impulso cardíaco
desde las aurículas hacia los ventrículos a través del haz AV.
La elevación de la concentración de
potasio hasta solo 8 a 12 mEq/l (dos a tres veces el valor normal) puede producir una profunda
debilidad del corazón, una alteración del ritmo e incluso la muerte.
Efecto de la temperatura sobre la función cardíaca

El aumento de la temperatura corporal, como ocurre durante la fiebre, produce un gran aumento de la frecuencia cardíaca, a veces hasta del doble del valor normal. El descenso de la temperatura produce una gran disminución de la frecuencia cardíaca, que puede disminuir hasta solo algunos latidos por minuto cuando una persona está cerca de la muerte por hipotermia en el intervalo de temperatura corporal de 16 °C a 21 °C.
La fuerza contráctil del corazón con frecuencia se incrementa transitoriamente cuando hay un aumento moderado de la temperatura, como ocurre durante el ejercicio corporal, aunque una elevación prolongada de la temperatura agota los sistemas metabólicos del corazón y finalmente produce debilidad.
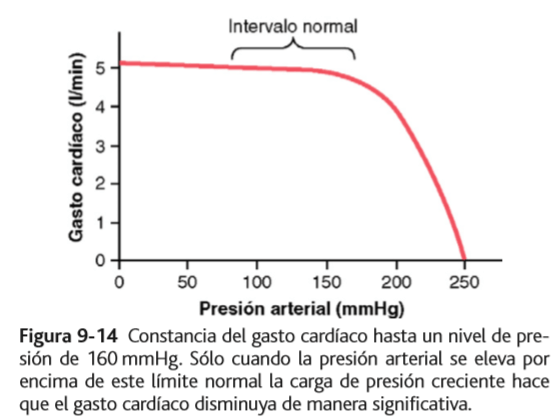
El incremento de la carga de presión arterial (hasta un límite)
no disminuye el gasto cardíaco
Durante la función normal del corazón a presiones arteriales sistólicas normales (80 a 140 mmHg) el gasto cardíaco está determinado casi totalmente por la facilidad del flujo sanguíneo a través de los tejidos corporales, que a su vez controla el retorno venoso de la sangre hacia el corazón
