El sistema urinario: anatomía funcional y
formación de orina en los riñones
Múltiples funciones del riñón en la homeostasis
Los riñones realizan sus funciones más importantes filtrando el plasma y eliminando sustancias del filtrado con una intensidad variable, dependiendo de las necesidades del cuerpo. Finalmente, los riñones «aclaran» las sustancias no deseadas del filtrado (y por tanto del cuerpo) excretándolas a la orina mientras devuelven las sustancias necesarias de nuevo a la sangre.

• Excreción de productos metabólicos de desecho y sustancias químicas extrañas.
Regulación de los equilibrios hídrico y electrolítico.
• Regulación de la osmolalidad del líquido corporal y de las concentraciones de electrólitos.

• Regulación de la presión arterial.
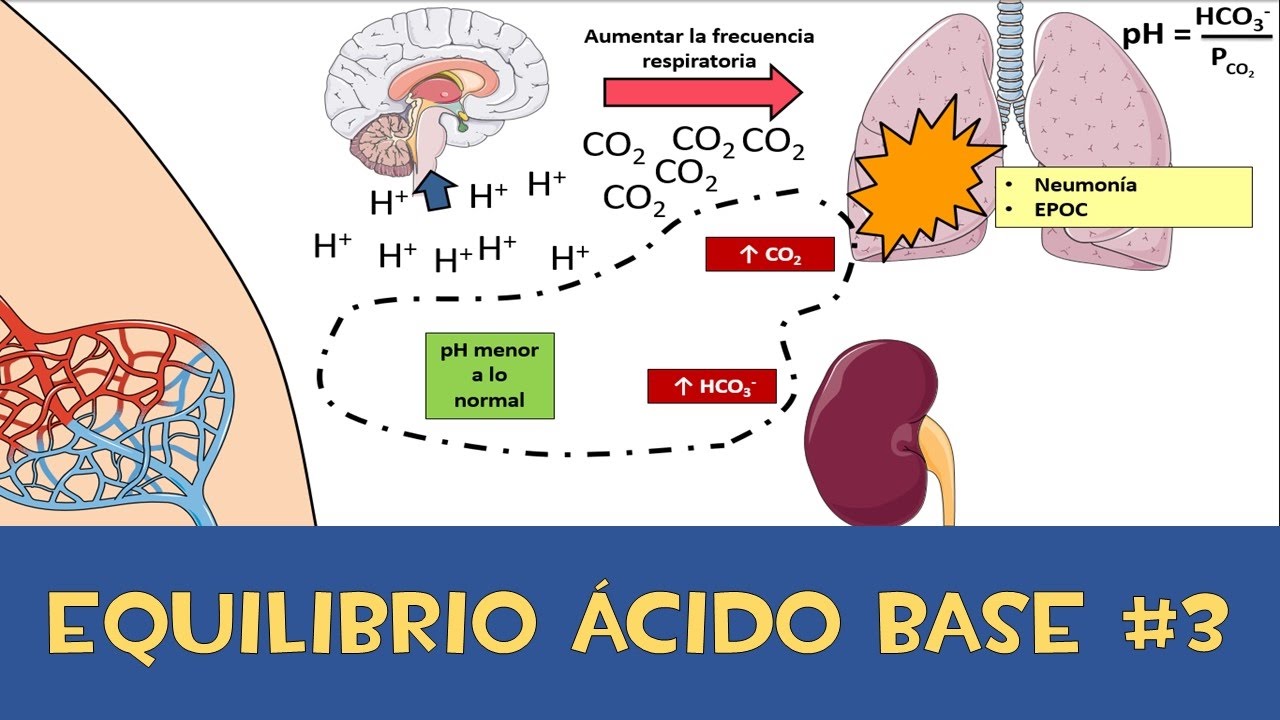
• Regulación del equilibrio acidobásico.

• Regulación de la producción de eritrocitos.
• Secreción, metabolismo y excreción de hormonas.
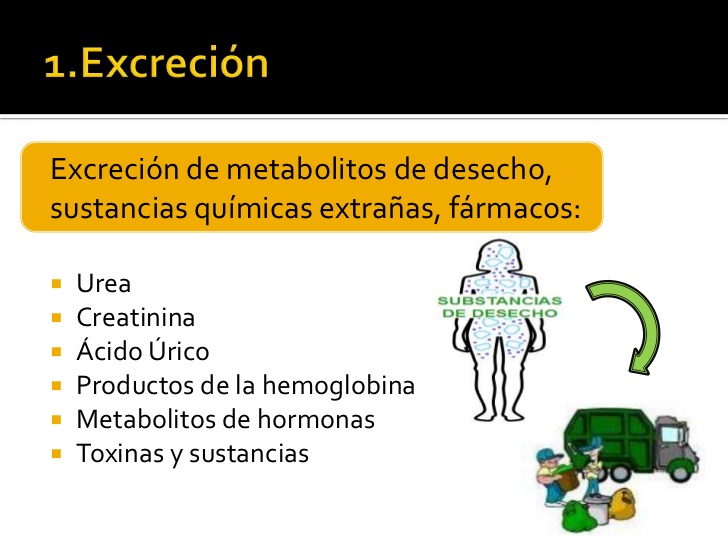
Excreción de productos metabólicos de desecho, sustancias químicas extrañas, fármacos y metabolitos de hormonas
Los riñones son los principales medios de eliminación de los productos de desecho del metabolismo que ya no necesita el cuerpo. Estos productos son la urea (del metabolismo de los aminoácidos), la creatinina (de la creatina muscular), el ácido úrico (de los ácidos nucleicos), los productos finales del metabolismo de la hemoglobina (como la bilirrubina) y los metabolitos de varias hormonas.

Regulación de los equilibrios hídrico y electrolítico
Para el mantenimiento de la homeostasis, la excreción de agua y electrólitos debe corresponderse de forma precisa con su ingreso. Si los ingresos superan a la excreción, la cantidad de esa sustancia en el cuerpo aumentará. Si la ingestión es menor que la excreción, la cantidad de esa sustancia en el cuerpo se reducirá.
La ingestión de agua y de muchos electrólitos está gobernada sobre todo por los hábitos de bebida y comida de la persona, y los riñones deben ajustar su excreción a su ingestión.
La capacidad de los riñones de alterar la excreción de sodio en respuesta a los cambios en su ingestión es enorme.
Regulación de la presión arterial
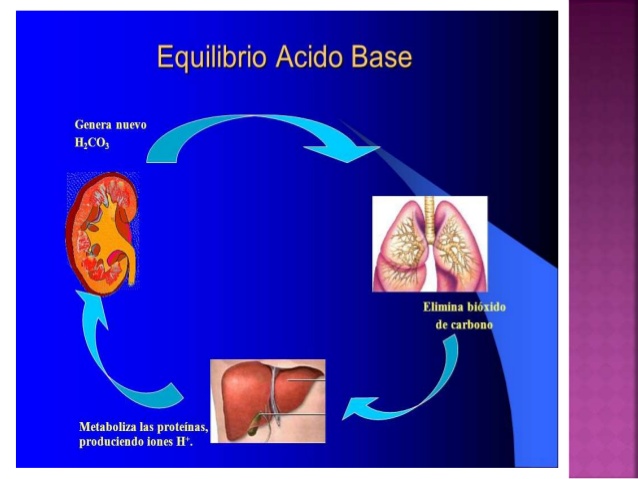
Los riñones contribuyen a la regulación acidobásica junto a los pulmones y los amortiguadores del líquido corporal mediante la excreción de ácidos y la regulación de los depósitos de amortiguadores en el líquido corporal.
Los riñones son el único medio de eliminar ciertos tipos de ácidos, como el
ácido sulfúrico y el ácido fosfórico, que genera el metabolismo de las proteínas.
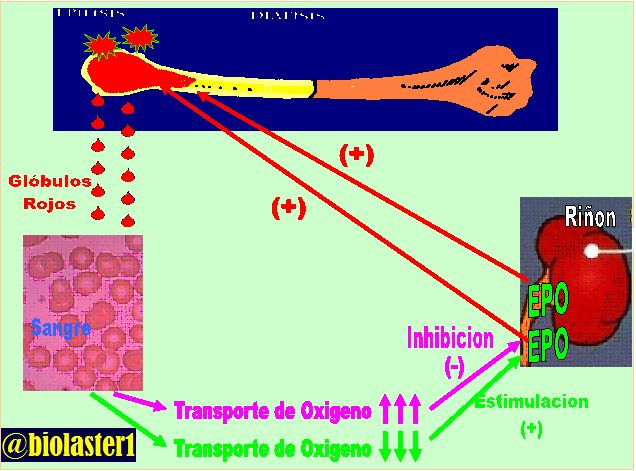
Regulación de la producción de eritrocitos
Los riñones secretan eritropoyetina, que estimula la producción de eritrocitos en las células madre hematopoyéticas de la médula ósea.
Los riñones son responsables normalmente de la mayor parte de la eritropoyetina secretada a la circulación. En las personas con una nefropatía grave o en las que se han extirpado los riñones y están en hemodiálisis, aparece una anemia grave debido a la menor producción de eritropoyetina.
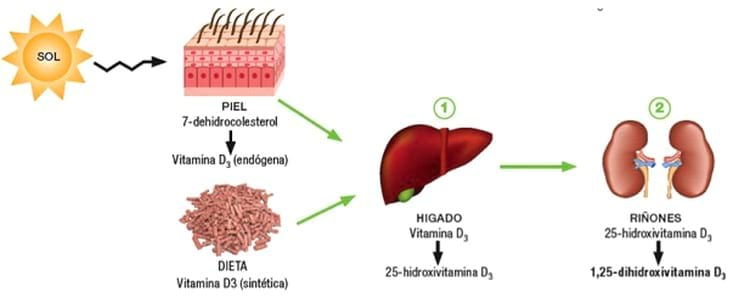
Regulación de la producción de 1,25-dihidroxivitamina D3
Los riñones producen la forma activa de la vitamina D, 1,25-dihidroxivitamina D3 (calcitriol), mediante la hidroxilación de esta vitamina en la posición «número 1».
El calcitriol es esencial para el depósito normal del calcio en el hueso y la reabsorción del calcio en el aparato digestivo
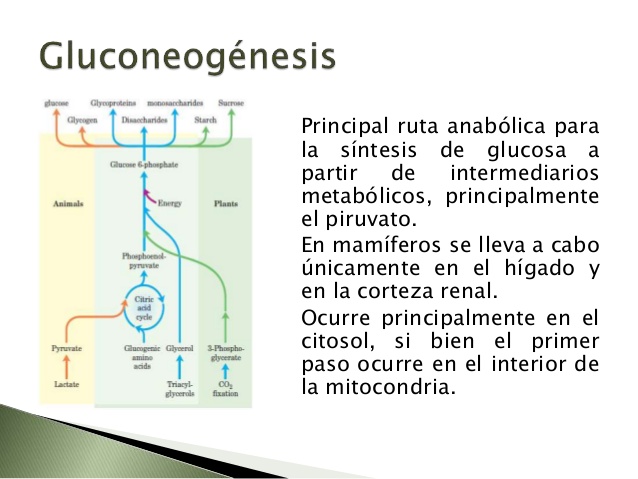
Síntesis de glucosa
Los riñones sintetizan glucosa a partir de los aminoácidos y otros precursores durante el ayuno prolongado, un proceso denominado gluconeogenia. La capacidad de los riñones de añadir glucosa a la sangre durante períodos prolongados de ayuno rivaliza con la del hígado.
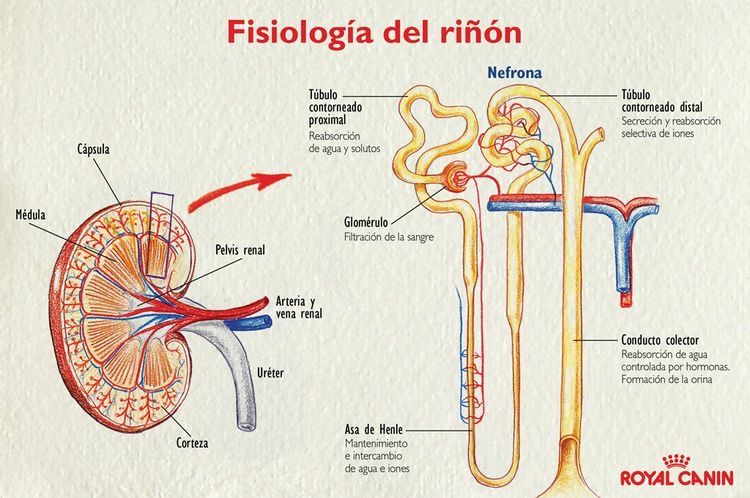
Anatomía fisiológica de los riñones
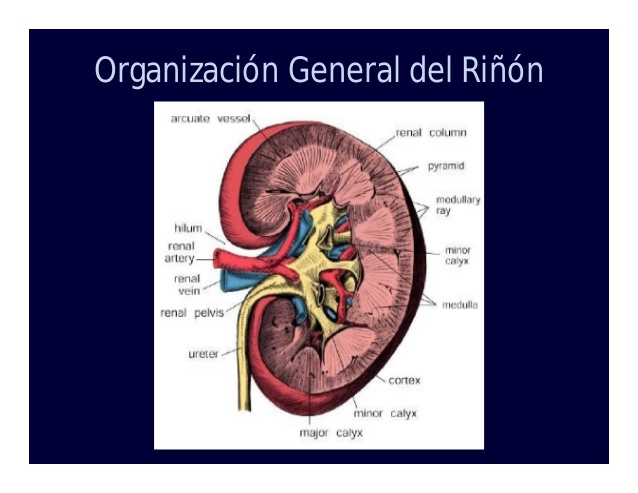
Organización general de los riñones y de la vía urinaria
Los riñones se disponen en la pared posterior del abdomen, fuera de la cavidad peritoneal.
Cada riñón de un ser humano adulto normal pesa unos 150 g y tiene el tamaño aproximado de unpuño cerrado.
La cara medial de cada riñón contiene una región con una muesca, llamada hilio, por la que pasan la arteria y vena renales, los linfáticos, la inervación y el uréter, que transporta la orina final desde el riñón hasta la vejiga, donde se almacena hasta que esta se vacía.
El riñón está rodeado de una cápsula fibrosa y tensa que protege sus delicadas estructuras internas.
La médula se divide en 8-10 masas de tejido en forma de cono llamadas pirámides renales.
La base de cada pirámide se origina en el borde entre la
corteza y la médula y termina en la papila, que se proyecta en el espacio de la pelvis renal, una continuación en forma de abanico de la porción superior del uréter.
El borde externo de la pelvis se divide en bolsas abiertas, llamadas cálices mayores, que se extienden hacia abajo y se dividen en los cálices menores, que recogen la orina de los túbulos de cada papila.
Las paredes de los cálices, la pelvis y el uréter contienen elementos contráctiles que empujan la orina hacia la vejiga, donde se almacena hasta que se vacía en la micción, que se comenta más adelante en este capítulo.
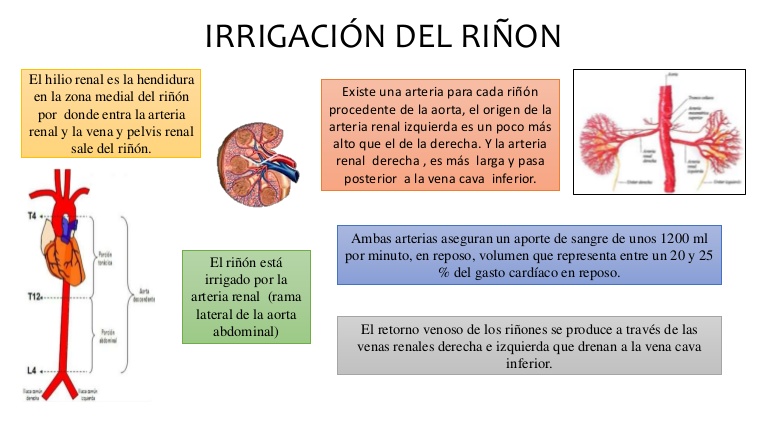
Irrigación renal
El riego sanguíneo de los dos riñones es normalmente de alrededor del 22% del gasto cardíaco, o 1.100 ml/min.
La arteria renal entra en el riñón a través del hilio y después se ramifica progresivamente hasta formar las arterias interlobulares, las arterias arciformes, las arterias interlobulillares (también denominadas arterias radiales) y las arteriolas aferentes, que acaban en los capilares glomerulares, donde se filtran grandes cantidades de líquido y solutos (excepto las proteínas
plasmáticas) para comenzar la formación de orina.
La circulación renal tiene la particularidad de contar con dos lechos capilares, los capilares glomerulares y los peritubulares, que están dispuestos en serie y están separados por las arteriolas eferentes.
Estas arteriolas ayudan a regular la presión hidrostática en los dos grupos de capilares. La presión hidrostática alta en los capilares glomerulares (de unos 60 mmHg) da lugar a una filtración rápida, mientras que una presión hidrostática mucho menor en los capilares peritubulares (de unos 13 mmHg) permite una reabsorción rápida de líquido.
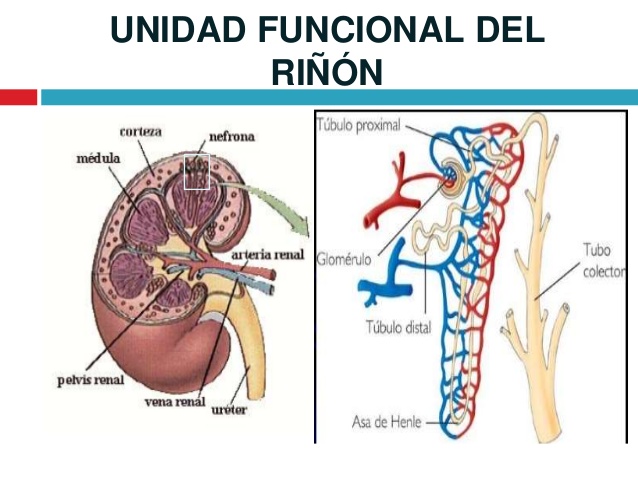
La nefrona es la unidad funcional del riñón
Cada riñón humano contiene alrededor de 800.000 a 1.000.000 de nefronas, cada una de las cuales es capaz de formar orina.
Cada nefrona contiene: 1) un penacho de capilares glomerulares llamado glomérulo, por el que se filtran grandes cantidades de líquido desde la sangre, y 2) un túbulo largo en el que el líquido filtrado se convierte en orina en su camino a la pelvis del riñón
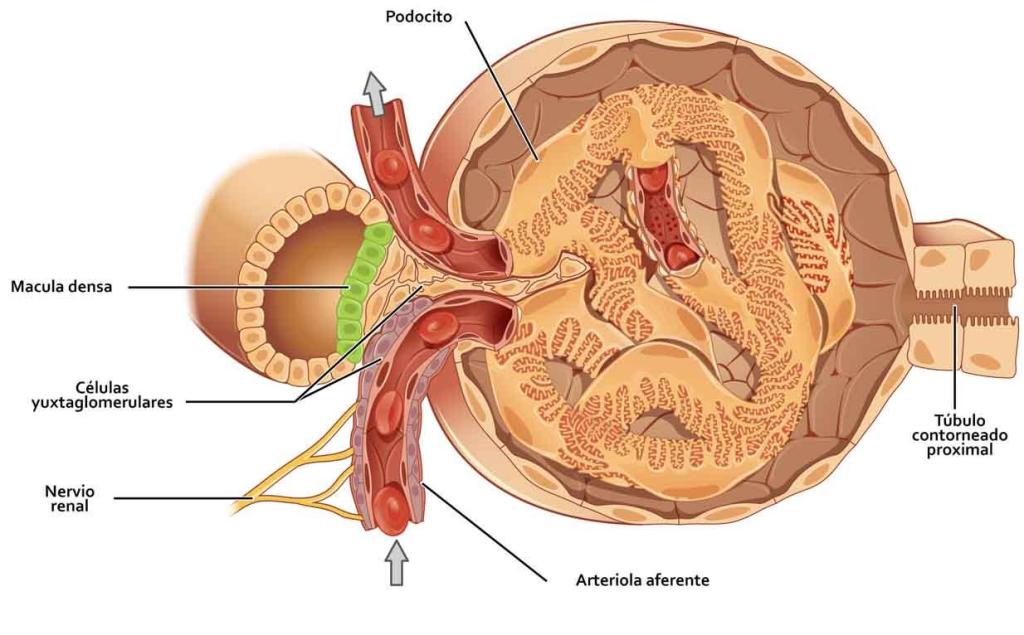
El glomérulo contiene una red de capilares glomerulares que se ramifican y anastomosan y que, comparados con otros capilares, tienen una presión hidrostática alta (de unos 60 mmHg). Los capilares glomerulares están revestidos de células epiteliales y todo el glomérulo está cubierto por la cápsula de Bowman.
El líquido filtrado desde los capilares glomerulares circula hacia la cápsula de Bowman y después al túbulo proximal, que se encuentra en la corteza del riñón
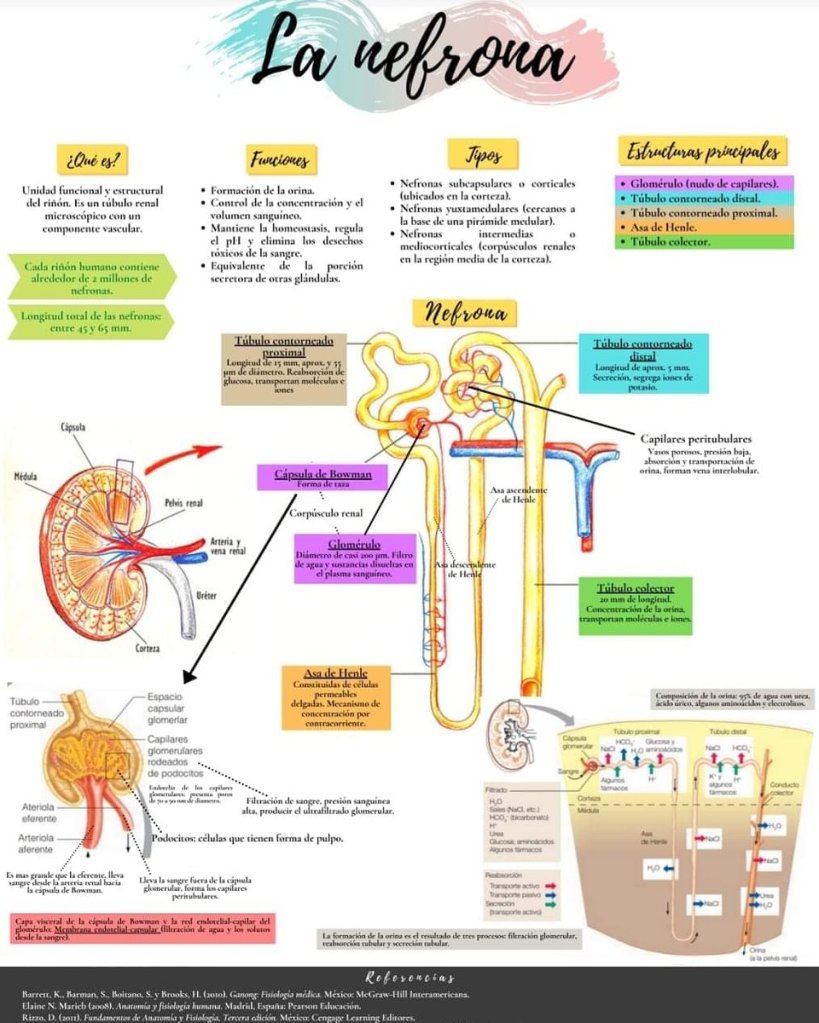
Al final de la rama ascendente gruesa hay un segmento corto, que tiene en su pared una placa de
células epiteliales especializadas conocida como mácula densa.
Más allá de la mácula densa el
líquido entra en el túbulo distal, que, como el túbulo proximal, se dispone en la corteza renal. Al túbulo distal le siguen el túbulo conector y el túbulo colector cortical, que conduce al conducto
colector cortical.
Las partes iniciales de 8 a 10 conductos colectores corticales se unen para formar un solo conducto colector mayor que discurre hacia abajo al interior de la médula y se convierte en el conducto colector medular. Los conductos colectores se funden para formar progresivamente conductos cada vez mayores que finalmente se vacían en la pelvis renal a través de las puntas de las
papilas renales.
Diferencias regionales en la estructura de la nefrona: nefronas corticales y
yuxtamedulares
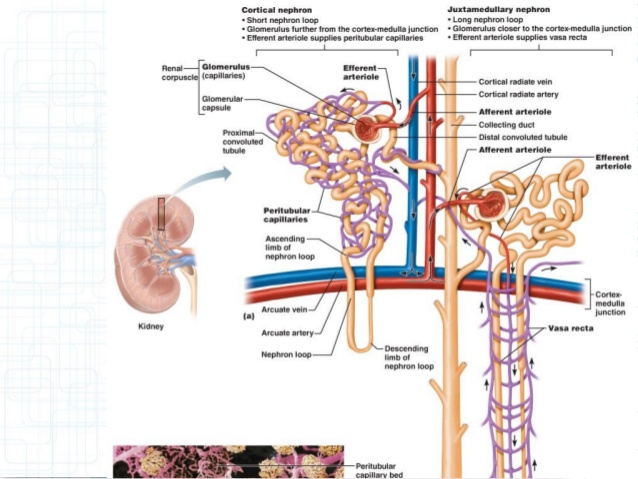
Nefronas corticales
Las nefronas que tienen
glomérulos localizados en la corteza externa se denominan nefronas corticales; tienen asas de Henle
cortas que penetran solo una distancia corta en la médula
Alrededor del 20-30% de las nefronas tienen glomérulos que se disponen en la profundidad de la
corteza renal cerca de la médula y se denominan nefronas yuxtamedulares. Estas nefronas tienen asas de Henle grandes que discurren hasta la médula, en algunos casos con un recorrido completamente intramedular hasta desembocar en las papilas renales.
Las estructuras vasculares que irrigan las nefronas yuxtamedulares también difieren de las que irrigan las nefronas corticales. En
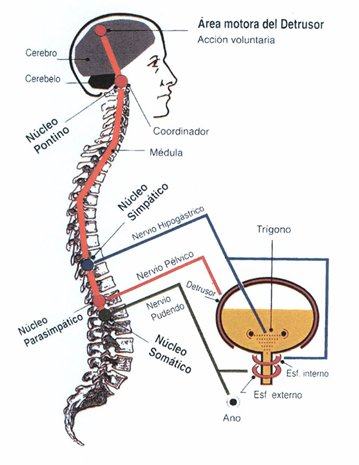
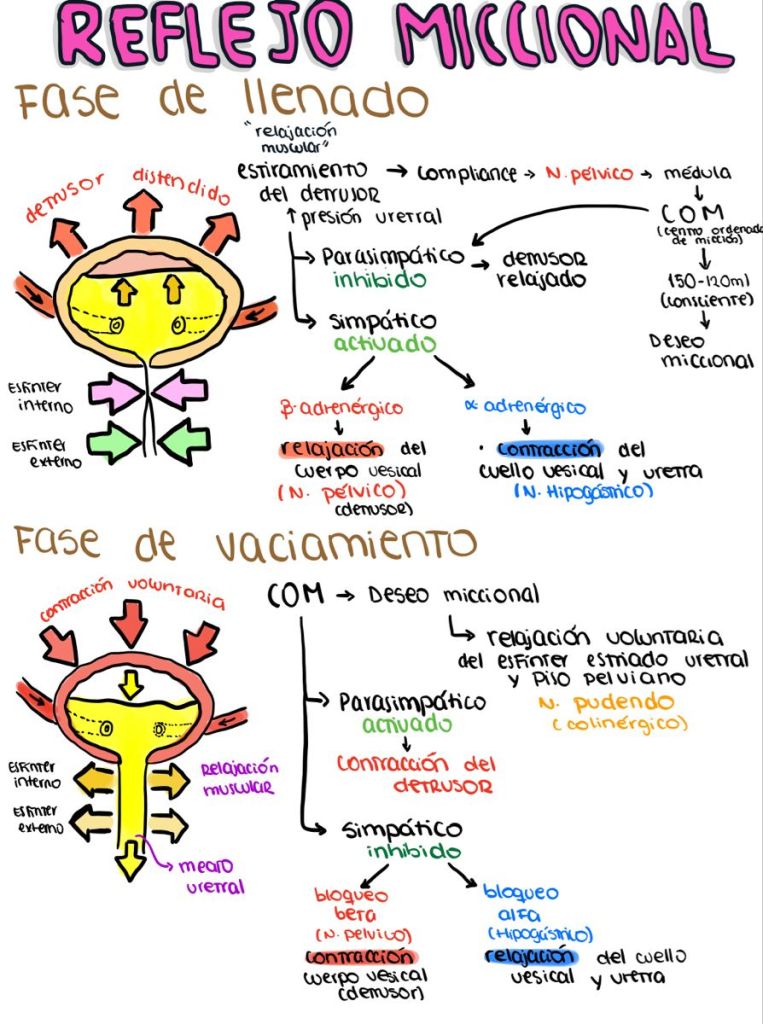
Micción
La micción es el proceso mediante el cual la vejiga urinaria se vacía cuando está llena.
Este proceso se realiza en dos pasos. Primero, la vejiga se llena progresivamente hasta que la tensión en sus
paredes aumenta por encima de un umbral. Esta tensión desencadena el segundo paso, que es un reflejo nervioso, llamado reflejo miccional, que vacía la vejiga o, si esto falla, provoca al menos un
deseo de orinar
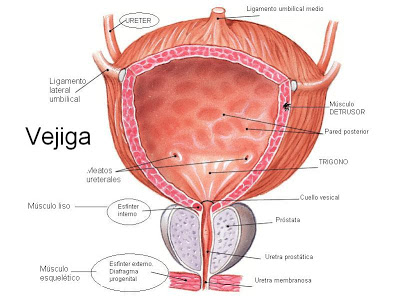
Anatomía fisiológica de la vejiga
Es una cámara de músculo liso compuesta de dos
partes principales:
- El cuerpo, que es la principal parte de la vejiga en la que se acumula la orina, y
- El cuello, que es una extensión en forma de abanico del cuerpo, que pasa en sentido inferior y anterior hasta el triángulo urogenital y se conecta con la uretra. La parte inferior del cuello de la vejiga también se llama uretra posterior por su relación con la uretra.
El músculo liso de la vejiga se llama músculo detrusor. Sus fibras musculares se extienden en todas las direcciones y, cuando se contraen, pueden aumentar la presión en la vejiga hasta 40-60 mmHg.
Luego la contracción del músculo detrusor es un paso importante en el vaciamiento de la vejiga. Las células musculares lisas del músculo detrusor se fusionan entre sí de manera que existen vías eléctricas de baja resistencia de una célula muscular a otra.
De este modo un potencial de acción puede propagarse a través del músculo detrusor, desde una célula muscular a la siguiente, para provocar la contracción de toda la vejiga a la vez.
Inervación de la vejiga

La principal inervación nerviosa de la vejiga es a través de los nervios pélvicos, que conectan con la médula espinal a través del plexo sacro, sobre todo los segmentos S2 y S3
En los nervios pélvicos discurren fibras nerviosas sensitivas y motoras. Las fibras sensitivas detectan el grado de distensión de la pared de la vejiga. Las señales de distensión de la uretra posterior son especialmente fuertes y son responsables sobre todo de iniciar los reflejos que provocan el vaciado de la vejiga.
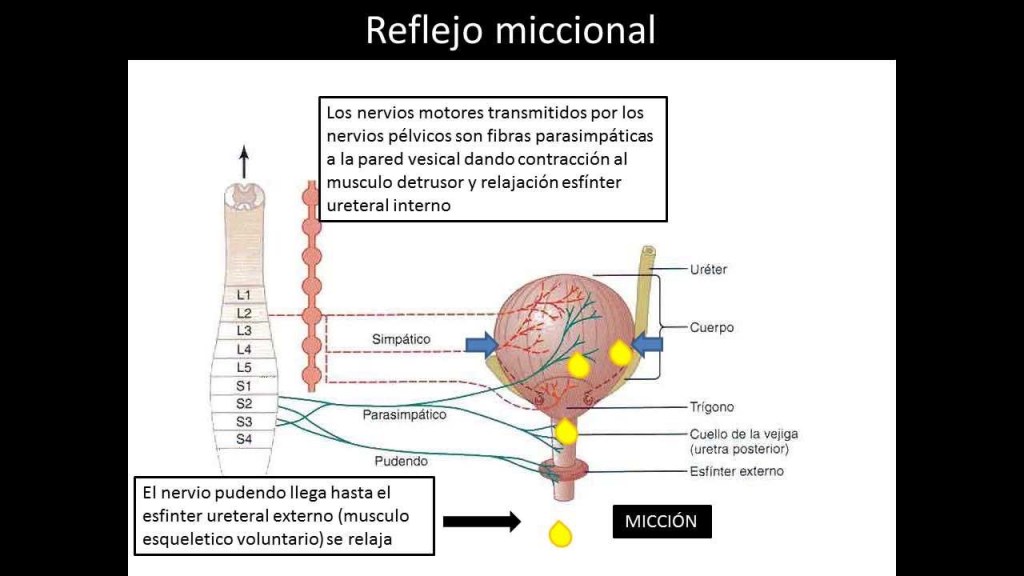
Los nervios motores transmitidos en los nervios pélvicos son fibras parasimpáticas.
Estas fibras terminan en las células ganglionares localizadas en la pared de la vejiga. Después, nervios posganglionares cortos inervan el músculo detrusor.
Además de los nervios pélvicos, otros dos tipos de inervación son importantes para la función vesical.
Los más importantes son las fibras motoras esqueléticas que llegan a través del nervio pudendo hasta el esfínter vesical externo. Se trata de fibras nerviosas somáticas que inervan ycontrolan el músculo esquelético voluntario del esfínter.
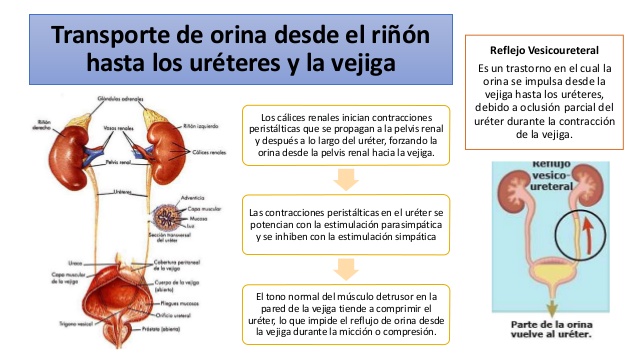
Transporte de orina desde el riñón hasta los uréteres y la
vejiga
La orina que sale de la vejiga tiene prácticamente la misma composición que el líquido que fluye de los conductos colectores; no hay cambios significativos en la composición de la orina en su camino a través de los cálices renales hasta los uréteres y la vejiga.
La orina que fluye desde los conductos colectores hacia los cálices renales estira los cálices e incrementa su actividad de marcapasos intrínseca, lo que a su vez inicia las contracciones peristálticas que se propagan a la pelvis renal y después a lo largo de la longitud del uréter, forzando así la orina desde la pelvis renal hacia la vejiga.
En los adultos, los uréteres tienen normalmente de 25 a 35 cm de longitud.
Las paredes de los uréteres contienen músculo liso y están inervadas por nervios simpáticos y parasimpáticos, así como por un plexo intramural de neuronas y fibras nerviosas que se extiende a lo largo de toda la longitud de los uréteres. Igual que sucede con otros músculos lisos viscerales, las contracciones peristálticas en el uréter se potencian con la estimulación parasimpática y se inhiben con la estimulación simpática.
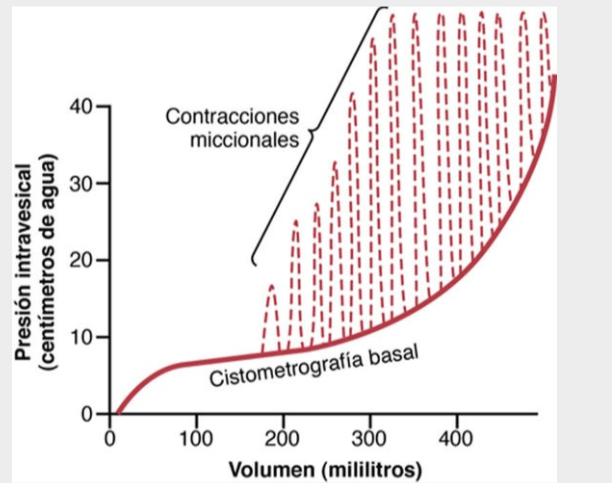
Llenado de la vejiga y tono de la pared vesical; la cistometrografía
Cuando no hay orina en la vejiga, la presión intravesical es aproximadamente de 0,
pero cuando se han acumulado 30-50 ml de orina, la presión aumenta a 5-10 cmH2O.
Puede recogerse una cantidad adicional (200-300 ml) de orina con solo pequeños aumentos adicionales de la presión; este nivel constante de presión se debe al tono intrínseco de la pared de la vejiga.
Por encima de los 300-400 ml, la acumulación de orina en la vejiga provoca un aumento rápido de la presión.
Reflejo miccional
El reflejo miccional es un solo ciclo completo de:
- Aumento rápido y progresivo de la presión
- Un período de presión mantenida
- Un retorno de la presión al tono basal de la vejiga.
Una vez que se ha producido el reflejo miccional pero no se ha vaciado la vejiga, los elementos nerviosos de este reflejo suelen permanecer en un estado de inhibición durante unos minutos a 1 h o más debido a que aparece otro reflejo miccional. A medida que la vejiga se llena má y más, los reflejos miccionales son más y más frecuentes y poderosos.
Una vez que el reflejo miccional es lo suficientemente poderoso, provoca otro reflejo, que pasa través de los nervios pudendos hasta el esfínter externo para inhibirlo. Si esta inhibición es má potente en el encéfalo que las señales constrictoras voluntarias al esfínter externo, se produce l micción.
Si no, la micción no se produce hasta que la vejiga se llena más y el reflejo miccional se hace más potente.
Facilitación o inhibición de la micción por el encéfalo

El reflejo miccional es un reflejo medular autónomo, pero centros encefálicos pueden inhibirlo o facilitarlo.
Estos centros son:
- Centros facilitadores e inhibidores potentes situados en el tronco del encéfalo, sobre todo en la protuberancia
- Varios centros localizados en la corteza cerebral que son sobre todo inhibidores, pero pueden hacerse excitadores.
- El reflejo miccional es la causa básica de la micción, pero los centros superiores ejercen normalmente un control final sobre la micción como sigue:
- Los centros superiores mantienen el reflejo miccional parcialmente inhibido, excepto cuando se desea la micción.
- Los centros superiores pueden impedir la micción, incluso aunque se produzca el reflejo miccional, mediante una contracción tónica del esfínter vesical externo hasta que se presente un momento adecuado.
- Cuando es el momento de la micción, los centros corticales pueden facilitar que los centros de la micción sacros ayuden a iniciar el reflejo miccional y al mismo tiempo inhibir el esfínter urinario externo para que la micción pueda tener lugar.
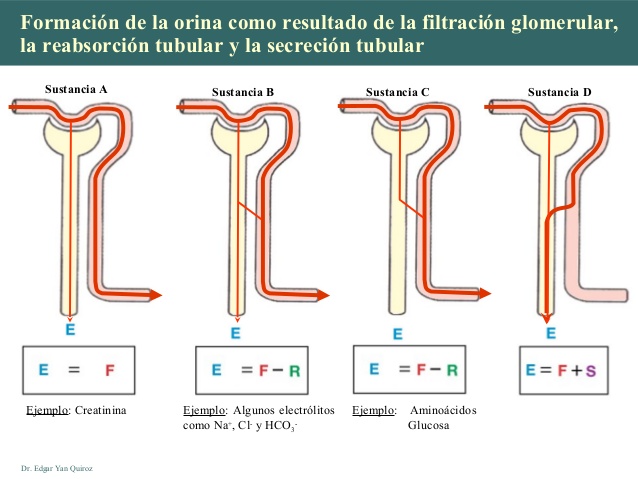
La formación de orina es resultado de la filtración
glomerular, la reabsorción tubular y la secreción
tubular
La intensidad con la que se excretan diferentes sustancias en la orina representa la suma de tres procesos renales
- La filtración glomerular
- La reabsorción de sustancias de los túbulos renales hacia la sangre
- La secreción de sustancias desde la sangre hacia los túbulos renales.
La formación de orina comienza cuando una gran cantidad de líquido que casi no dispone de proteínas se filtra desde los capilares glomerulares a la cápsula de Bowman. La mayoría de las sustancias del plasma, excepto las proteínas, se filtran libremente, de manera que su concentración en el filtrado glomerular de la cápsula de Bowman es casi la misma que en el plasma.
A medida que el líquido abandona la cápsula de Bowman y pasa a través de los túbulos, se modifica por la reabsorción de agua y solutos específicos de nuevo hacia la sangre o por la secreción de otras sustancias desde los capilares peritubulares hacia los túbulos.
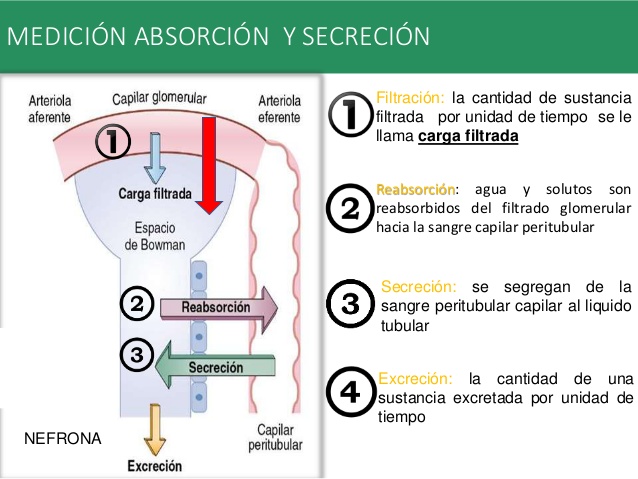
Filtración, reabsorción y secreción de diferentes sustancias
En general, la reabsorción tubular es cuantitativamente más importante que la secreción tubular en la formación de la orina, pero la secreción es importante para determinar las cantidades de iones potasio e hidrógeno y algunas otras sustancias que se excretan por la orina.
La mayoría de las sustancias que deben eliminarse de la sangre, en especial los productos finales del metabolismo, como la urea, la creatinina, el ácido úrico y los uratos, se reabsorben mal y por ello se excretan en grandes cantidades en la orina. Ciertas sustancias extrañas y fármacos se reabsorben mal pero, además, se secretan desde la sangre a los túbulos, de manera que su excreción es alta.
Por el contrario, los electrólitos, como los iones cloro, sodio y bicarbonato, se reabsorben mucho, de manera que solo se detectan en la orina pequeñas cantidades. Ciertas sustancias nutritivas, como los aminoácidos y la glucosa, se reabsorben completamente de los túbulos y no aparecen en la orina, aunque los capilares glomerulares filtren grandes cantidades.
