Hormonas corticosuprarrenales
Las dos glándulas suprarrenales, con un peso aproximado de 4 g cada una, se hallan en los polos superiores de los riñones.
La médula suprarrenal, que ocupa el 20% central de la glándula, se relaciona desde el punto de vista funcional con el sistema nervioso simpático; secreta las hormonas adrenalina y noradrenalina en respuesta a la estimulación simpática.
A su vez, estas hormonas provocan casi los mismos efectos que la estimulación directa de los nervios simpáticos en todas las regiones del cuerpo.
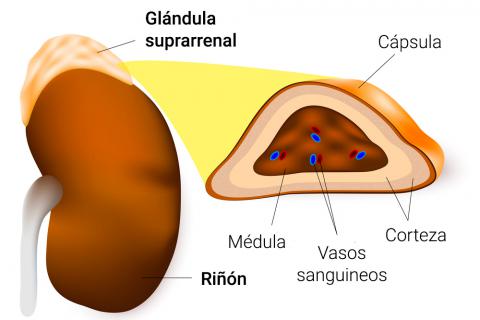
La corteza suprarrenal secreta un grupo completamente diferente de hormonas, llamadas corticoesteroides.
Todas estas hormonas se sintetizan a partir del esteroide colesterol y todas poseen
una fórmula química parecida. Sin embargo, las pequeñas variaciones de su estructura molecular proporcionan diferencias funcionales muy importantes.
Corticoesteroides: mineralocorticoides,
glucocorticoides y andrógenos
La corteza suprarrenal secreta los dos tipos principales de hormonas corticosuprarrenales, los mineralocorticoides y los glucocorticoides. Además de estas hormonas, produce pequeñas cantidades de hormonas sexuales, en particular de andrógenos, que inducen los mismos efectos que la hormona sexual masculina testosterona.
Los mineralocorticoides reciben este nombre porque afectan sobre todo a los electrólitos (los
«minerales») del compartimiento extracelular, especialmente al sodio y al potasio. Los glucocorticoides se denominan así porque poseen efectos importantes de aumento de la glucemia.
Además, influyen en el metabolismo de las proteínas y de los lípidos, con efectos tan importantes para la función del organismo como los que producen sobre el metabolismo de los hidratos de carbono.
Síntesis y secreción de hormonas
corticosuprarrenales
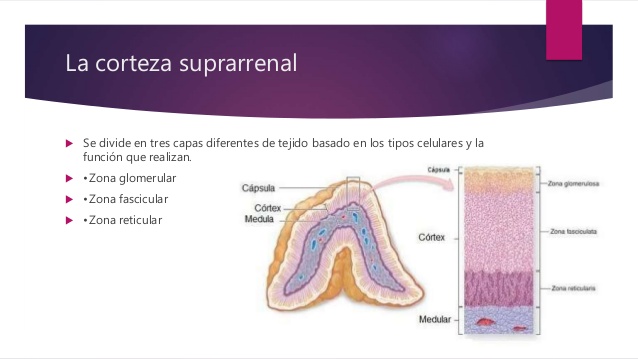
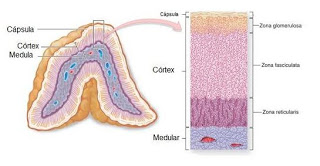
La corteza suprarrenal tiene tres capas diferentes
Contribuye con casi el 15% a la corteza suprarrenal. Estas células son las únicas de la glándula suprarrenal capaces de secretar cantidades importantes de aldosterona porque contienen la enzima aldosterona sintetasa, necesaria para la síntesis de la hormona.
La secreción de estas células está controlada sobre todo por las concentraciones de angiotensina II y potasio en el líquido extracelular ambos estimulan la secreción de aldosterona.
- La zona fascicular, la zona media y más ancha, representa casi el 75% de la corteza suprarrenal y secreta los glucocorticoides cortisol y corticosterona, así como pequeñas cantidades de andrógenos y estrógenos suprarrenales. La secreción de estas células está controlada, en gran parte, por el eje
hipotalámico-hipofisario a través de la corticotropina (ACTH). - La zona reticular, la capa más profunda de la corteza, secreta los andrógenos suprarrenales deshidroepiandrosterona (DHEA) y androstenodiona, así como pequeñas cantidades de estrógenos y algunos glucocorticoides. La ACTH también regula la secreción de estas células, aunque en ella pueden intervenir otros factores tales como la hormona corticótropa estimuladora de los andrógenos, liberada por la hipófisis. Sin embargo, los mecanismos que regulan la producción suprarrenal de andrógenos no se conocen tan bien como los de los glucocorticoides y mineralocorticoides.
Vías de síntesis de los esteroides suprarrenales
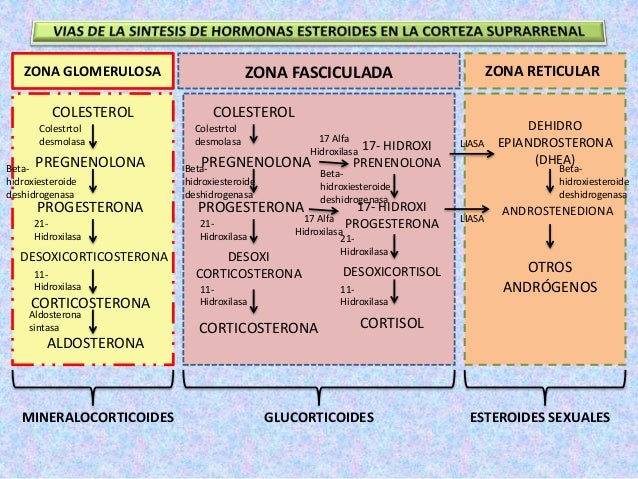
Cada etapa está catalizada por un sistema enzimático específico.
Un cambio, incluso de una sola enzima, puede provocar la formación de tipos muy distintos y porcentajes diferentes de hormonas. Por ejemplo, si se altera la actividad de tan solo una enzima de esta vía, se generarán cantidades enormes de hormonas sexuales masculinizantes u otros compuestos esteroideos que normalmente no se encuentran en la sangre.
Las fórmulas químicas de la aldosterona y el cortisol, que son las principales hormonas mineralocorticoide y glucocorticoide, respectivamente
Además de la aldosterona y del cortisol, la corteza suprarrenal suele secretar pequeñas cantidades de otros esteroides con actividad glucocorticoide, mineralocorticoide o mixta. Por último, se han sintetizado y empleado en diversas formas de tratamiento varias hormonas esteroideas potentes, no sintetizadas por las glándulas suprarrenales en condiciones normales.
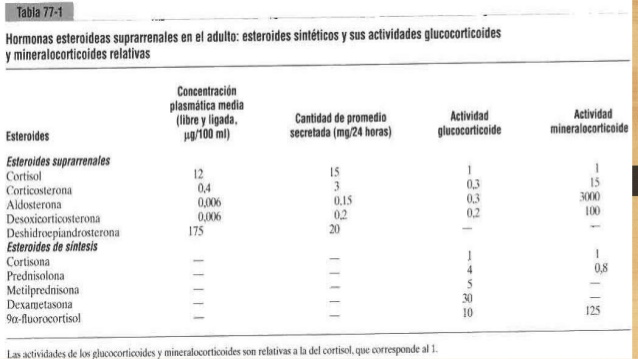
Hormonas esteroideas suprarrenales en el adulto
• Aldosterona (muy potente, supone casi el 90% de toda la actividad mineralocorticoide).
• Desoxicorticosterona (1/30 de la potencia de la aldosterona, aunque se secreta en cantidades
mínimas).
• Corticosterona (ligera actividad mineralocorticoide).
• 9α-fluorocortisol (sintético, algo más potente que la aldosterona).
• Cortisol (actividad mineralocorticoide mínima, pero se secreta en grandes cantidades).
• Cortisona (actividad mineralocorticoide mínima).

• Cortisol (muy potente; es el responsable de casi el 95% de toda la actividad glucocorticoide).
• Corticosterona (proporciona el 4% de la actividad glucocorticoide total, pero es mucho menos
potente que el cortisol).
• Cortisona (casi tan potente como el cortisol).
• Prednisona (sintética, cuatro veces más potente que el cortisol).
• Metilprednisolona (sintética, cinco veces más potente que el cortisol).
• Dexametasona (sintética, 30 veces más potente que el cortisol).
De la lista anterior se desprende que algunas de estas hormonas y
Las hormonas corticosuprarrenales se unen a las proteínas del plasma
Aproximadamente del 90 al 95% del cortisol plasmático se une a las proteínas del plasma, sobre todo a una globulina denominada globulina fijadora del cortisol o transcortina, y en menor grado, a la albúmina.
Esta unión tan fuerte a las proteínas del plasma reduce la velocidad de eliminación del cortisol plasmático; por tanto, el cortisol posee una semivida relativamente larga, de 60 a 90 min.
Tan solo el 60% de la aldosterona circulante se une a las proteínas del plasma, de modo que el 40% queda en forma libre; en consecuencia, su semivida es bastante reducida, de unos 20 min.
Funciones de los mineralocorticoides: aldosterona

La deficiencia de mineralocorticoides provoca pérdidas renales intensas de cloruro
sódico e hiperpotasemia
La pérdida completa de la secreción corticosuprarrenal puede causar la muerte en un plazo de 3 días a 2 semanas, salvo que la persona reciba un tratamiento salino intensivo o la inyección de mineralocorticoides.
Sin mineralocorticoides, la concentración del ion potasio del líquido extracelular experimenta un gran ascenso, el sodio y el cloruro desaparecen enseguida del organismo y el volumen total del líquido extracelular y el volumen de sangre se reducen mucho.
Pronto se desarrolla un descenso del gasto cardíaco, que evoluciona a un estado de shock, seguido de la muerte.
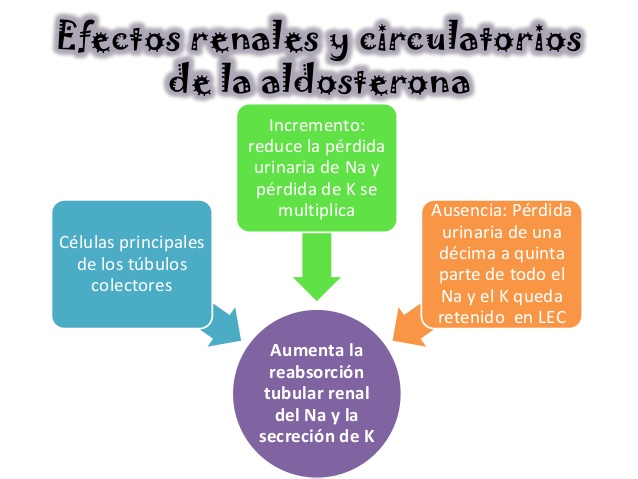
Efectos renales y circulatorios de la aldosterona
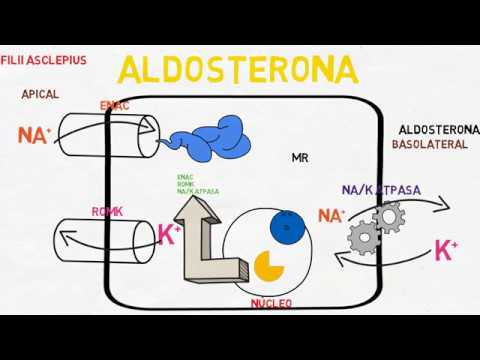
La aldosterona aumenta la reabsorción tubular renal del sodio y la secreción de potasio
El incremento de la concentración de aldosterona del plasma puede reducir de forma pasajera la pérdida urinaria de sodio y dejarla en unos pocos miliequivalentes al día. Al mismo tiempo, las pérdidas de potasio por la orina se multiplican transitoriamente. Así pues, el efecto neto del exceso de aldosterona en el plasma consiste en un aumento de la cantidad total de sodio en el líquido extracelular y un descenso de la de potasio.
Por el contrario, la ausencia total de secreción de aldosterona puede ocasionar una pérdida urinaria pasajera de 10 a 20 g de sodio al día, cantidad equivalente a una décima a quinta parte de todo el sodio orgánico. Al mismo tiempo, el potasio queda retenido con tenacidad en el líquido extracelular.
El exceso de aldosterona produce hipopotasemia y debilidad muscular; el déficit de
aldosterona induce hiperpotasemia y toxicidad cardíaca
El exceso de aldosterona no solo provoca la de iones potasio desde el líquido extracelular hacia la orina, sino que también estimula el transporte de potasio desde el líquido extracelular hacia la mayoría de las células del organismo.
Por consiguiente, la secreción excesiva de aldosterona, como ocurre con algunos tipos de tumores suprarrenales, induce un descenso importante de la concentración plasmática de potasio, a veces desde valores normales de 4,5 mEq/l hasta cifras tan bajas como 2 mEq/l. Esta situación se denomina hipopotasemia

La aldosterona estimula el transporte de sodio y potasio en
las glándulas sudoríparas, las glándulas salivales y las
células epiteliales intestinales
La aldosterona ejerce casi los mismos efectos sobre las glándulas sudoríparas y salivales que sobre los túbulos renales. Estos dos tipos de glándulas producen una secreción primaria que contiene grandes cantidades de cloruro sódico, aunque gran parte del cloruro sódico se reabsorbe al atravesar los conductos excretores, mientras que los iones potasio y bicarbonato se excretan.
La aldosterona aumenta de manera considerable la reabsorción de cloruro sódico y la secreción de potasio por los conductos.
El efecto sobre las glándulas sudoríparas reviste interés para conservar la sal del
organismo en ambientes cálidos y el efecto sobre las glándulas salivales permite conservar la sal cuando se pierden cantidades excesivas de saliva.
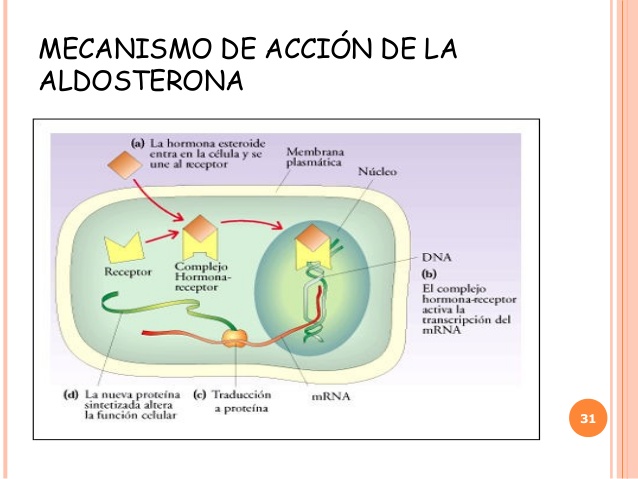
Mecanismo celular de la acción de la aldosterona
Desde hace muchos años se conocen los efectos generales de los mineralocorticoides sobre el organismo, pero se ignora el mecanismo molecular de la acción de la aldosterona incrementa el transporte de sodio en las células tubulares. No obstante, la secuencia celular de acontecimientos que culmina con el aumento de la reabsorción de sodio parece ser la siguiente.
En primer lugar, la aldosterona difunde de inmediato al interior de las células del epitelio tubular, debido a su liposolubilidad en las membranas celulares

Posibles acciones no genómicas de la aldosterona y otras
hormonas esteroideas
Algunos estudios indican que muchos esteroides, entre ellos la aldosterona, no solo producen efectos genómicos de desarrollo lento, con una latencia de 45 a 60 min y que dependen de la transcripción del gen y de la síntesis de nuevas proteínas, sino que también tienen efectos más rápidos no genómicos, que aparecen luego de tan solo pocos segundos a algunos minutos.
Parece que estas acciones no genómicas dependen de la unión de los esteroides a receptores de la membrana celular que se acoplan a sistemas de segundos mensajeros, similares a los utilizados por las hormonas peptídicas para la transmisión de señales.
Regulación de la secreción de aldosterona
La regulación de la secreción de aldosterona por las células de la zona glomerular no depende apenas de la regulación del cortisol o de los andrógenos por las zonas fascicular y reticular.
Se conocen los siguientes cuatro factores que desempeñan una función esencial para la regulación de la aldosterona:
- El incremento de la concentración de iones potasio en el líquido extracelular aumenta mucho la secreción de aldosterona.
- El aumento de la concentración de angiotensina II en el líquido extracelular también incrementa mucho la secreción de aldosterona.
- El incremento de la concentración de iones sodio en el líquido extracelular apenas reduce la secreción de aldosterona.
- Se necesita ACTH de la adenohipófisis para que haya secreción de aldosterona, aunque su efecto regulador sobre la velocidad de secreción es mínimo en la mayoría de los trastornos fisiológicos.

Funciones de los glucocorticoides.
Los mineralocorticoides pueden salvar la vida de los animales sometidos a suprarrenalectomía aguda, pero estos animales no se encuentran ni mucho menos bien. En realidad, los sistemas metabólicos animales de utilización de las proteínas, hidratos de carbono y lípidos están muy alterados.
Además, el animal no resiste ningún tipo de estrés físico o mental y cualquier enfermedad leve, como una infección respiratoria, puede causar su muerte.
Efectos del cortisol sobre el metabolismo de los hidratos de carbono

Estimulación de la gluconeogenia
El efecto metabólico más conocido del cortisol y de otros glucocorticoides consiste en estimular la gluconeogenia (es decir, la formación de hidratos de carbono a partir de las proteínas y de otras sustancias) en el hígado; el ritmo de gluconeogenia se eleva, a menudo, entre 6 y 10 veces.
Este aumento del ritmo de la gluconeogenia se debe, sobre todo, a los efectos directos del cortisol en el hígado, así como a la antagonización de los efectos de la insulina.
- El cortisol aumenta las enzimas que convierten los aminoácidos en glucosa dentro de los hepatocitos. Este efecto se debe a la capacidad de los glucocorticoides para activar la transcripción del ADN en el núcleo del hepatocito, de la misma manera que la aldosterona actúa en las células del
túbulo renal: se forman ARNm que, a su vez, dan origen al conjunto de las enzimas necesarias para la gluconeogenia. - El cortisol moviliza los aminoácidos de los tejidos extrahepáticos, sobre todo del músculo. Por ello, llegan más aminoácidos al plasma, para incorporarse a la gluconeogenia hepática y facilitar la formación de glucosa.
- El cortisol antagoniza los efectos de la insulina para inhibir la gluconeogenia en el hígado.
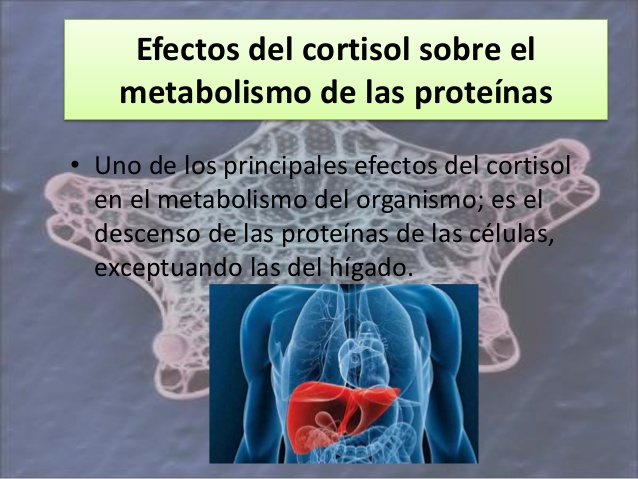
Efectos del cortisol sobre el metabolismo de las proteínas
Uno de los principales efectos del cortisol sobre los sistemas metabólicos del organismo consiste en el descenso de los depósitos de proteínas de la práctica totalidad de las células del organismo, con excepción de las del hígado. Esta reducción se debe tanto al descenso de la síntesis como a un mayor
catabolismo de las proteínas ya existentes dentro de las células.
Ambos efectos podrían achacarse a un menor transporte de los aminoácidos a los tejidos extrahepáticos, como se expondrá más adelante; es posible que esta no sea la causa primordial, porque el cortisol también reduce la formación de
ARN y la síntesis posterior de proteínas de muchos tejidos extrahepáticos, sobre todo del músculo y del tejido linfático.
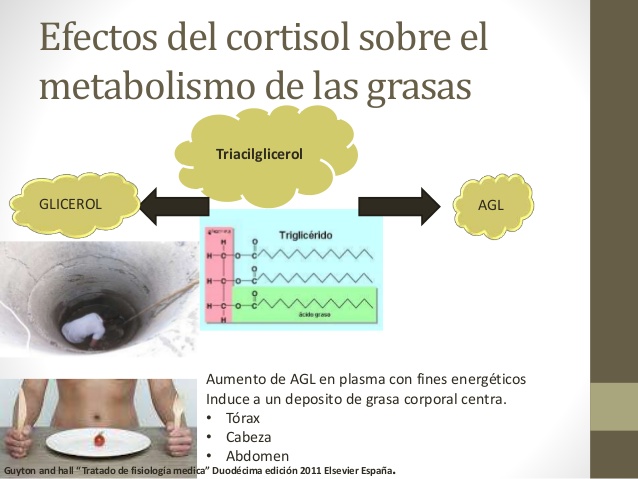
Efectos del cortisol sobre el metabolismo de las grasas
De forma muy similar a la movilización de aminoácidos del músculo, el cortisol moviliza a los ácidos grasos del tejido adiposo. Esta movilización aumenta la concentración de ácidos grasos libres en el plasma, lo que aumenta también la utilización de los ácidos grasos con fines energéticos. Al parecer, el cortisol ejerce asimismo un efecto directo que potencia la oxidación de los ácidos grasos en el interior de la célula.
En los períodos de ayuno prolongado o de estrés, la mayor movilización de grasas por el cortisol, junto con el incremento en la oxidación de los ácidos grasos en la célula, inducen una desviación de los sistemas metabólicos celulares, que pasan de la utilización energética de glucosa a la utilización de ácidos grasos.
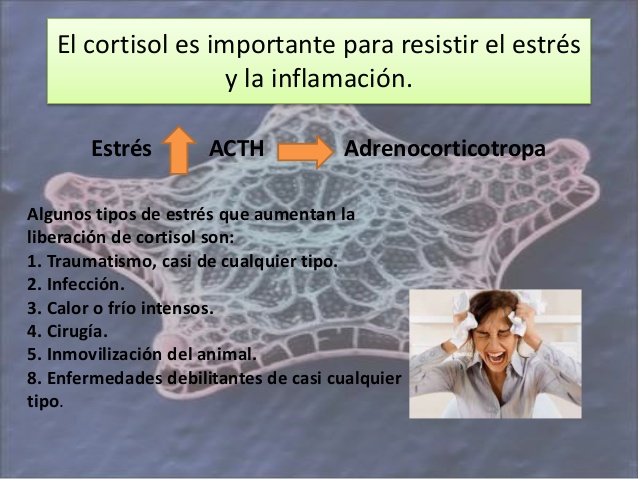
El cortisol es importante para resistir el estrés y la inflamación
Prácticamente cualquier tipo de estrés, ya sea físico o neurógeno, provoca un aumento inmediato y notable de la secreción de ACTH por la adenohipófisis, seguido unos minutos después de una secreción considerable de cortisol por la corteza suprarrenal.
Efectos antiinflamatorios de las concentraciones altas de cortisol
Cuando un tejido sufre daños a causa de un traumatismo, una infección bacteriana o cualquier otra causa, suele «inflamarse».
A veces, como ocurre en la artritis reumatoide, la inflamación resulta más nociva que el traumatismo o la enfermedad. La administración de grandes cantidades de cortisol permite, de ordinario, bloquear esta inflamación o incluso revertir muchos de sus efectos, una vez iniciada. Antes de explicar los mecanismos por los que el cortisol bloquea la inflamación
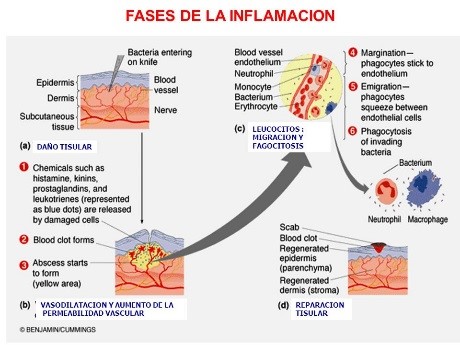
La inflamación consta de cinco etapas fundamentales
- Liberación por las células dañadas del tejido de sustancias químicas que activan el proceso inflamatorio, tales como histamina, bradicinina, enzimas proteolíticas, prostaglandinas y leucotrienos
- Aumento del flujo sanguíneo en la zona inflamada, inducido por alguno de los productos liberados de los tejidos, un efecto que se denomina eritema
- Salida de grandes cantidades de plasma casi puro desde los capilares hacia las zonas dañadas, secundaria a un aumento de la permeabilidad capilar, seguida de la coagulación del líquido tisular, con el consiguiente edema sin fóvea
- Infiltración de la zona por leucocitos
- Crecimiento de tejido fibroso pasados unos días o semanas, para contribuir a la cicatrización.
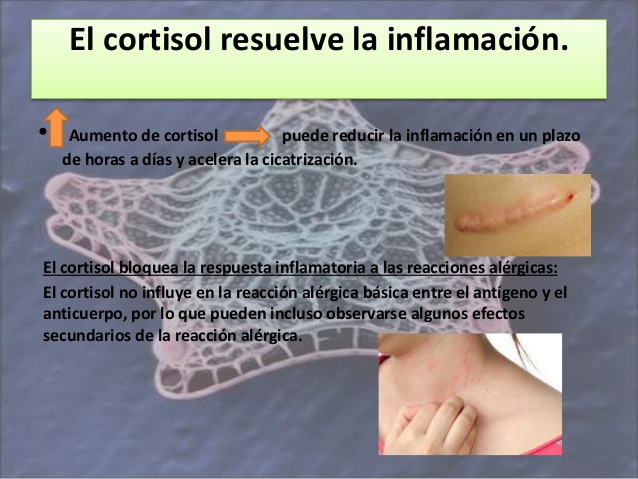
El cortisol resuelve la inflamación
Incluso después de establecida la inflamación, la administración de cortisol puede reducirla en un plazo de horas a días.
Bloquea casi todos los factores que fomentan el proceso y, además, acelera la
cicatrización. Sin embargo, es muy probable que la explicación resida en los mismos factores, en su mayoría desconocidos, que permiten al organismo resistir muchos otros tipos de estrés físico cuando se secretan grandes cantidades de cortisol.
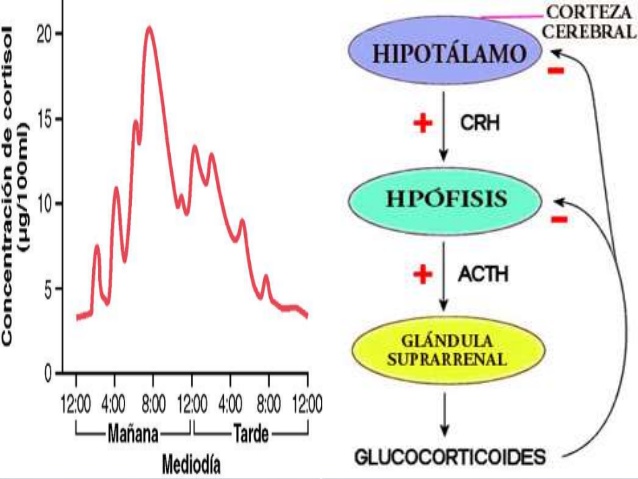
Regulación de la secreción de cortisol por la corticotropina
procedente de la hipófisis

A diferencia de la secreción de aldosterona en la zona glomerular, controlada sobre todo por el
potasio y la angiotensina II que actúan directamente sobre las células de la corteza suprarrenal, la secreción de cortisol está sometida de forma casi exclusiva al control de la ACTH hipofisaria.

La ACTH se ha aislado de forma pura de la adenohipófisis. Es un polipéptido grande, correspondiente a una cadena de 39 aminoácidos. Se conoce un polipéptido menor, un producto de la
digestión de la ACTH, cuya cadena mide 24 aminoácidos pero que posee todos los efectos de la
molécula entera.
La corticoliberina hipotalámica controla la secreción de ACTH:
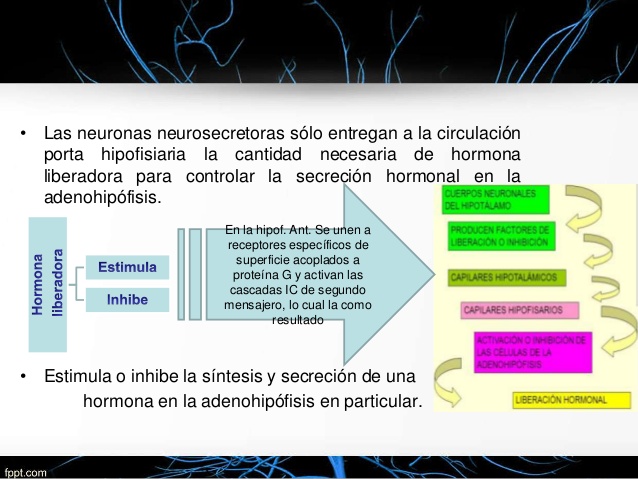
De manera idéntica al control de otras hormonas hipofisarias por los factores liberadores del
hipotálamo, un factor liberador importante controla la liberación de ACTH. Se llama corticoliberina
o factor liberador de corticotropina (CRF). Se secreta hacia el plexo capilar primario del sistema
hipofisario portal en la eminencia media del hipotálamo y luego se transporta a la adenohipófisis,
donde induce la secreción de ACTH. El CRF es un péptido formado por 41 aminoácidos
Síntesis y secreción de ACTH asociadas a las de hormona
estimuladora de los melanocitos, lipotropina y endorfina
Cuando la adenohipófisis secreta ACTH, al mismo tiempo se liberan otras hormonas de estructura química parecida. Ello se debe a que el gen se transcribe para formar la molécula de ARN que determina la formación de ACTH y provoca, en principio, la síntesis de una proteína mucho mayor,una preprohormona llamada proopiomelanocortina (POMC) que, además de ser la precursora de la
ACTH, también da lugar a otros varios péptidos, entre los que se encuentran la hormona estimuladora de los melanocitos (MSH), la β-lipotropina además de otros y la β-endorfina.

En los abundantes melanocitos localizados entre la dermis y la epidermis de la piel, la MSH
estimula la formación de un pigmento negro, la melanina, que se dispersa hacia la epidermis. La
inyección de MSH a una persona a lo largo de 8 a 10 días oscurece mucho la piel.
La ACTH, que contiene una secuencia de la MSH, ejerce un efecto estimulante de los melanocitos que equivale a alrededor de 1/30 del de la MSH. Además, como las cantidades de MSH pura secretada por el ser humano son muy reducidas y las de ACTH, abundantes, es probable que la ACTH tenga más importancia que la MSH para establecer la cantidad de melanina de la piel.
Anomalías de la secreción corticosuprarrenal

Hipofunción corticosuprarrenal (insuficiencia corticosuprarrenal):
enfermedad de Addison
La enfermedad de Addison se debe a la incapacidad de la corteza suprarrenal para fabricar suficientes hormonas corticales; a su vez, en un elevado número de casos, la causa obedece a una atrofia o lesión primaria de la corteza suprarrenal. Esta atrofia se debe casi en el 80% de las ocasiones a un fenómeno de autoinmunidad dirigido contra la corteza suprarrenal.
La hipofunción de las glándulas suprarrenales puede ocurrir también por destrucción tuberculosa o por la invasión de la corteza por un tumor maligno.
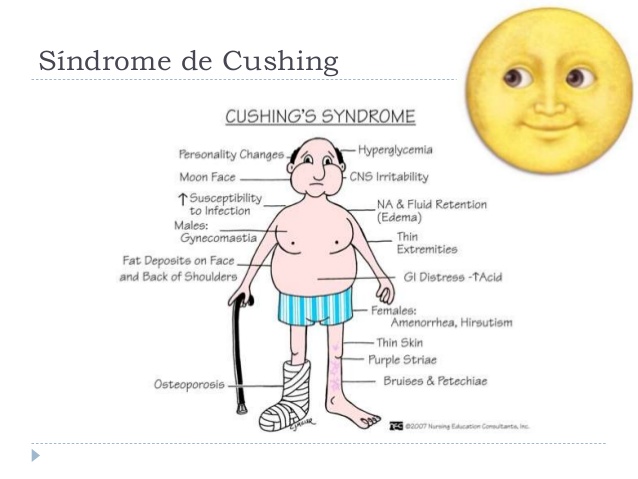
Hiperfunción corticosuprarrenal: síndrome de Cushing
La hipersecreción corticosuprarrenal provoca una cascada compleja de efectos hormonales, conocida como síndrome de Cushing.
Muchas anomalías de este síndrome se deben al exceso de cortisol, aunque la secreción exagerada de andrógenos también ocasiona efectos importantes.
El hipercortisolismo obedece a múltiples causas, por ejemplo:
- Adenomas adenohipofisarios secretores de grandes cantidades de ACTH que, a su vez, causan hiperplasia suprarrenal y exceso de cortisol
- Anomalías de la función del hipotálamo que ocasionan un aumento de liberación de la hormona liberadora de corticotropina, con el consiguiente estímulo exagerado de la secreción de ACTH
- Secreción ectópica de ACTH por un tumor de otra parte del cuerpo, como un carcinoma abdominal
- Adenomas de la corteza suprarrenal.
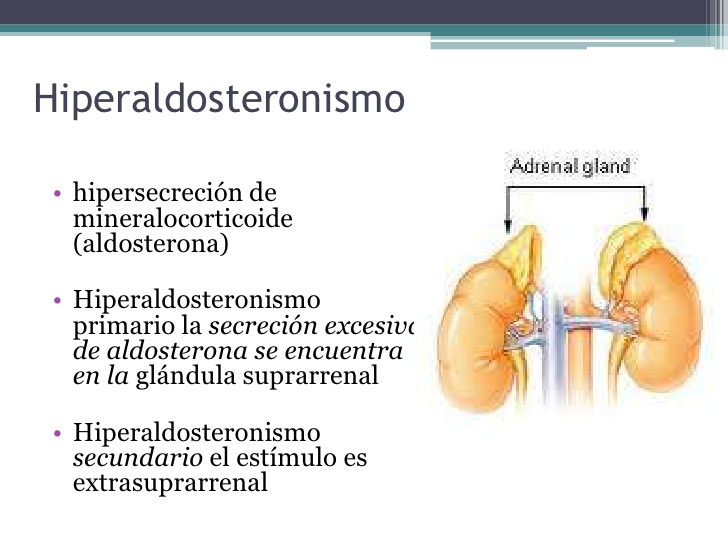
Hiperaldosteronismo primario (síndrome de Conn)
A veces se desarrolla un pequeño tumor en la zona glomerular y se produce una gran secreción de aldosterona; el estado resultante se conoce como hiperaldosteronismo primario o síndrome de Conn.
Asimismo, en casos aislados, la corteza suprarrenal hiperplásica secreta aldosterona y no cortisol.
Los efectos del exceso de aldosterona se expusieron ya con detalle en este mismo capítulo.
Las consecuencias más importantes son hipopotasemia, alcalosis metabólica leve, un ligero aumento del volumen extracelular y del volumen sanguíneo, un incremento mínimo de la concentración plasmática de sodio (normalmente <4 a 6 mEq/l) y, casi siempre, hipertensión.
Merecen destacarse, dentro del hiperaldosteronismo primario, los períodos ocasionales de parálisis muscular inducidos por la hipopotasemia.
