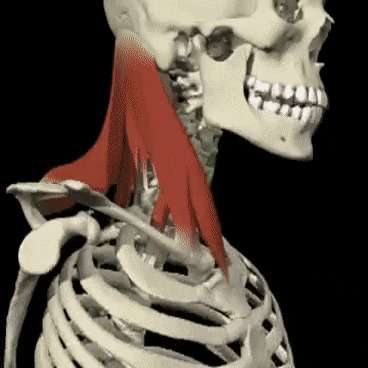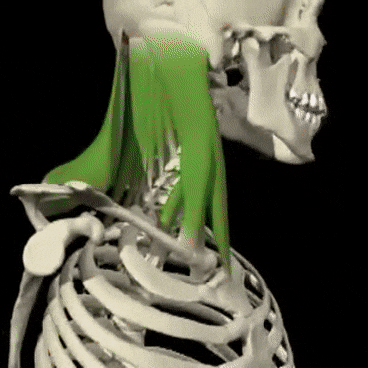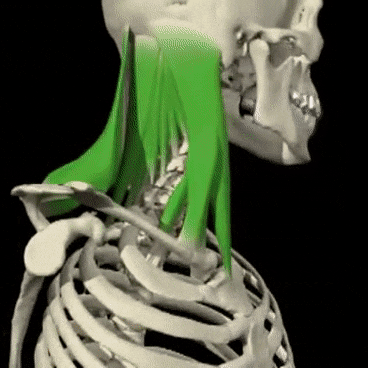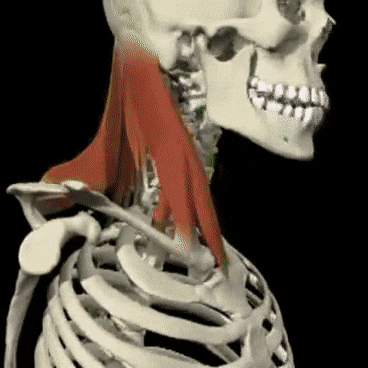
Mecanismos encefálicos del comportamiento y la motivación: el sistema límbico y el hipotálamo
Las señales nerviosas del tronco del encéfalo activan el componente cerebral del encéfalo por dos caminos:
- 1) mediante la estimulación directa de un nivel de actividad neuronal de fondo en amplias regiones del cerebro
- 2) por medio de la puesta en marcha de sistemas neurohormonales capaces de liberar neurotransmisores específicos facilitadores o inhibidores de tipo hormonal en determinadas zonas del encéfalo.
El sistema general encargado de controlar el nivel de actividad del encéfalo, su componente impulsor central consiste en una zona excitadora situada en la formación reticular de la protuberancia y el mesencéfalo denominación de área facilitadora bulborreticular
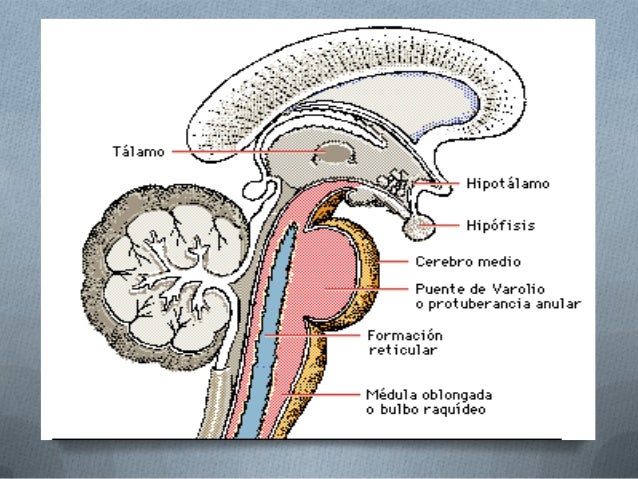
Además de señales descendentes, esta región también envía una abundancia de señales en sentido ascendente.
La mayoría de estas señales primero van al tálamo, donde excitan a un nuevo grupo de neuronas que transmiten señales nerviosas hacia todas las regiones de la corteza cerebral
Las señales que atraviesan el tálamo son de dos tipos:
- Uno consiste en unos potenciales de acción de conducción rápida que excitan el cerebro tan solo durante unos pocos milisegundos. Nacen en los grandes somas neuronales situados por toda el área reticular del tronco del encéfalo.
- El segundo tipo de señal excitadora se origina en una gran cantidad de pequeñas neuronas dispersas por todo el área reticular excitadora del tronco del encéfalo. Una vez más, la mayoría de ellas se dirigen hacia el tálamo, pero esta vez a través de pequeñas fibras de conducción lenta que hacen sinapsis sobre todo en los núcleos talámicos intralaminares
El nivel de actividad del área excitadora en el tronco del encéfalo y, por tanto, el de todo el encéfalo, viene determinado en gran medida por la cantidad y el tipo de las señales sensitivas que llegan al encéfalo desde la periferia.

A la corteza cerebral no solo llegan impulsos activadores desde el área excitadora bulborreticular del tronco del encéfalo, sino que también regresan señales de retroalimentación desde la corteza cerebral a esta misma área.
Se trata de un mecanismo general de retroalimentación positiva que permite un refuerzo aún mayor de la actividad con cualquier otra actividad iniciada en la corteza cerebral, lo que se traduce en una mente «despierta».
Ademas la estimulación eléctrica de un punto concreto en el tálamo en general activa su propia región particular restringida en la corteza. Además, por lo común las señales reverberan de un lado a otro entre el tálamo y la corteza cerebral, de modo que el primero excita a esta última y ella a continuación reexcita al tálamo a través de sus fibras de regreso.

Area inhibidora reticular: ocupa una posición medial y ventral en el bulbo raquídeo; tiene la capacidad para inhibir el área facilitadora reticular de la parte alta del tronco del encéfalo, y reducir así también la actividad de las porciones superiores del cerebro. Uno de los mecanismos seguidos para cumplir esta actividad consiste en excitar las neuronas serotoninérgicas, que a su vez segregan la neurohormona inhibidora serotonina en puntos cruciales del encéfalo
Control neurohormonal de la actividad encefálica:
control directo de la actividad cerebral efectuado mediante la transmisión específica de señales nerviosas desde las zonas inferiores del encéfalo hacia sus regiones corticales
segregar sustancias hormonales neurotransmisoras excitadoras o inhibidoras sobre el parénquima del encéfalo
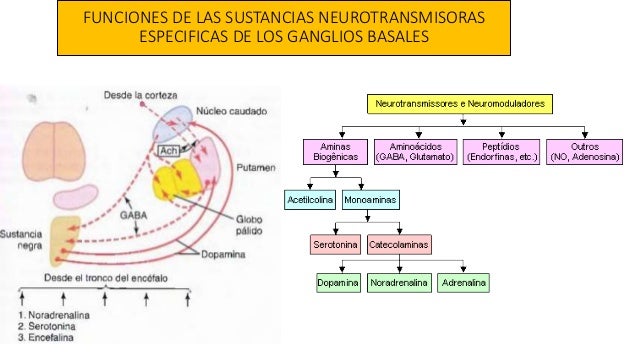
- 1) un sistema noradrenérgico: (suele actuar como una hormona excitador, este sistema está diseminado prácticamente por cualquiera de sus zonas)
- 2) un sistema dopaminérgico (dopamina: es excitadora en algunas zonas, pero inhibidora en otras, se dirige sobre todo a las áreas de los ganglios basales)
- 3) un sistema serotoninérgico.(serotonina normalmente posee un carácter inhibidor, se dirige especialmente hacia las estructuras de la línea media.)
Regiones del tronco del encéfalo humano donde se activan cuatro sistemas neurohormonales, los tres comentados para la rata y otro más, el sistema acetilcolinérgico.
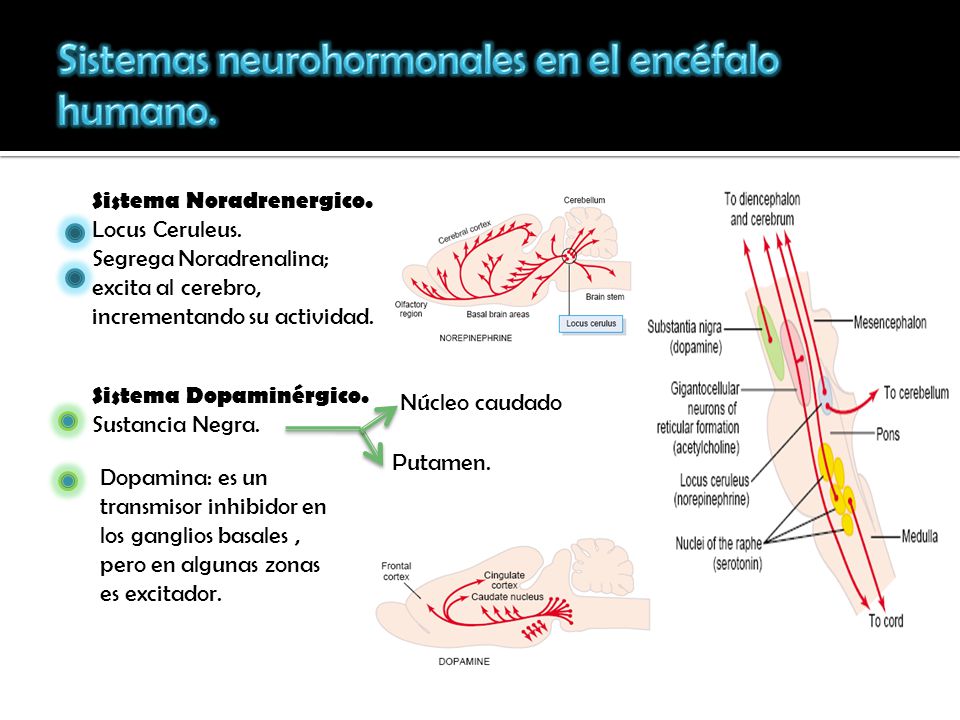
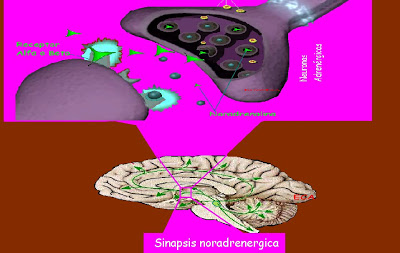
- El locus ceruleus y el sistema de la noradrenalina. El locus ceruleus es una pequeña zona que ocupa una posición bilateral y posterior en la unión entre la protuberancia y el mesencéfalo.Esta sustancia en general excita el encéfalo con el fin de incrementar su actividad. Sin embargo, posee unos efectos inhibidores en unas cuantas de sus regiones debido a los receptores de este tipo que existen en ciertas sinapsis neuronales.
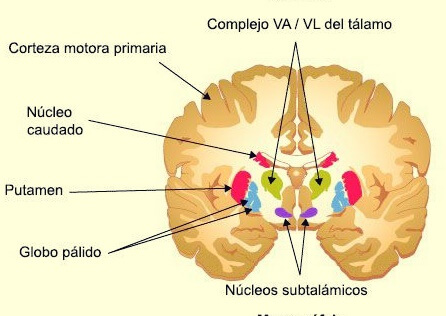
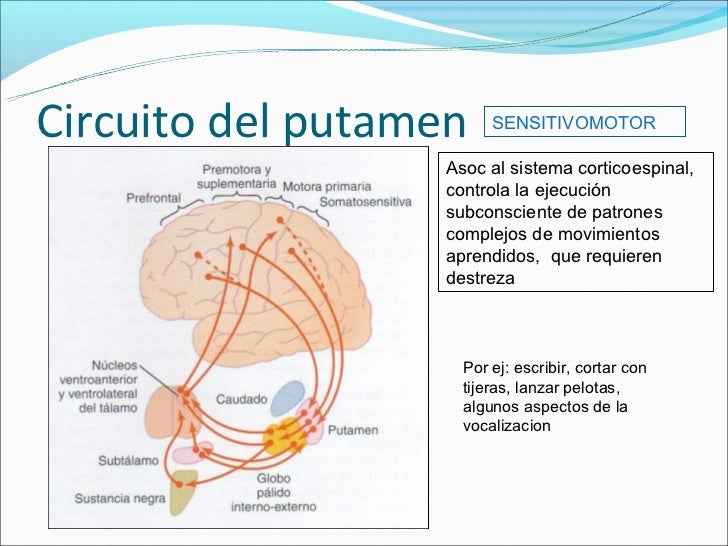
- La sustancia negra y el sistema de la dopamina: Ocupa una posición anterior en la parte superior del mesencéfalo, y sus neuronas envían terminaciones nerviosas sobre todo hacia el núcleo caudado y el putamen en el cerebro, donde segregan dopamina

- Los núcleos del rafe y el sistema de la serotonina. En la línea media de la protuberancia y el bulbo raquídeo hay varias estructuras delgadas llamadas núcleos del rafe. Muchas de las neuronas que les componen segregan serotonina. Envían sus fibras hacia el diencéfalo y unas cuantas hacia la corteza cerebral; aún otras más descienden hacia la médula espinal

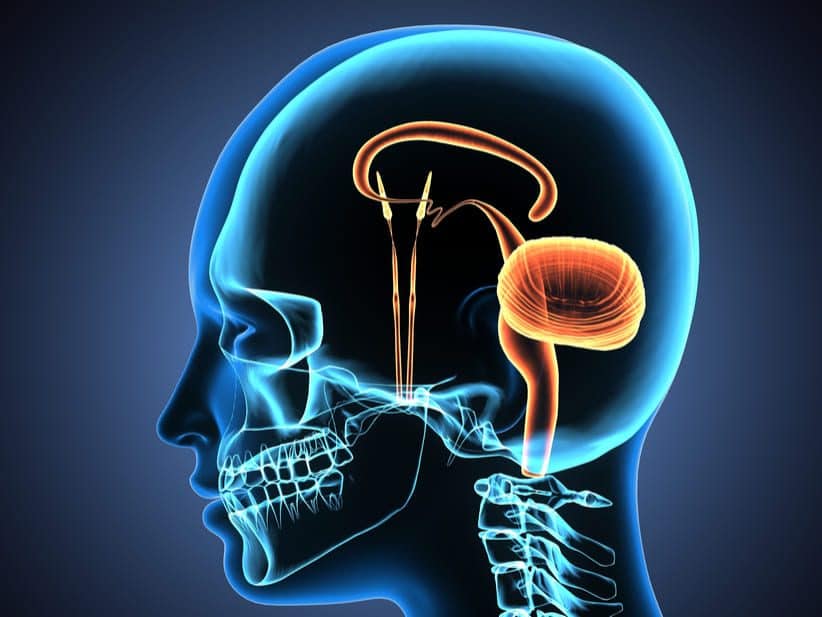
La palabra «límbico» significa «limítrofe». En su origen, este término se empleó para describir las estructuras fronterizas que rodean a las regiones basales del cerebro,
Circuito neuronal que controla el comportamiento emocional y los impulsos de las motivaciones.
Un componente fundamental del sistema límbico es el hipotálamo, regulan muchos estados internos del cuerpo, como:
- temperatura corporal
- la osmolalidad de los líquidos corporales
- impulsos para comer y beber
- controlar el peso corporal.



Estas funciones internas se denominan en su conjunto funciones vegetativas del encéfalo, y su control se encuentra íntimamente emparentado con el del comportamiento.
Sistema límbico es un un complejo interconectado de elementos basales del encéfalo.Situado en el centro de todas estas estructuras está el pequeñísimo hipotálamo, que desde un punto de vista fisiológico es uno de los componentes nucleares del sistema límbico.
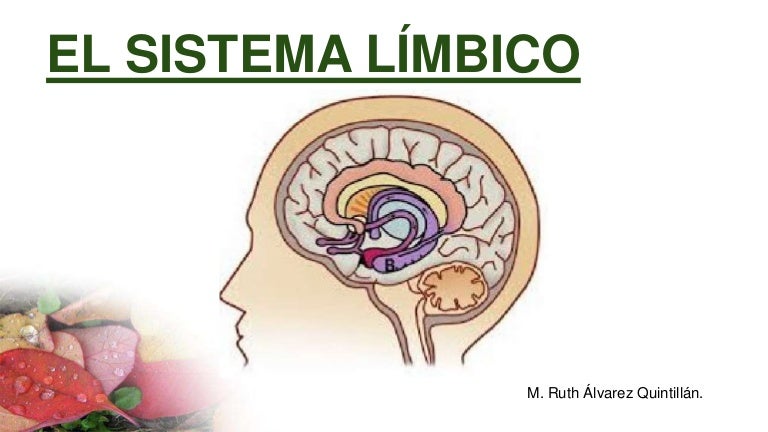
Las estructuras subcorticales pertenecientes a este sistema, entre ellas los núcleos septales, el área paraolfatoria, los núcleos anteriores del tálamo, ciertas porciones de los ganglios basales, el hipocampo y la amígdala.
En torno a las regiones límbicas subcorticales queda la corteza límbica, integrada por un anillo de corteza cerebral a cada lado del encéfalo:
- 1) que comienza en el área orbitofrontal de la cara ventral de los lóbulos frontales
- 2) asciende hacia la circunvolución subcallosa
- 3) a continuación sigue por encima de la parte superior del cuerpo calloso sobre la cara medial del hemisferio cerebral en la circunvolución cingular; y, finalmente
- 4) pasa por detrás del cuerpo calloso y desciende sobre la cara ventromedial del lóbulo temporal hacia la circunvolución parahipocámpica y el uncus.
Un camino importante de comunicación entre el sistema límbico y el tronco del encéfalo es el fascículo prosencefálico medial.

El hipotálamo posee vías de comunicación de doble sentido con todos los estratos del sistema límbico. Envíando señales eferentes en tres direcciones:
- 1) posterior e inferior, hacia el tronco del encéfalo, dirigidas sobre todo a las áreas reticulares del mesencéfalo, la protuberancia y el bulbo raquídeo, y desde estas regiones hacia los nervios periféricos pertenecientes al sistema nervioso autónomo
- 2) superior, hacia muchas zonas altas del diencéfalo y el telencéfalo, especialmente los núcleos anteriores del tálamo y las porciones límbicas de la corteza cerebral
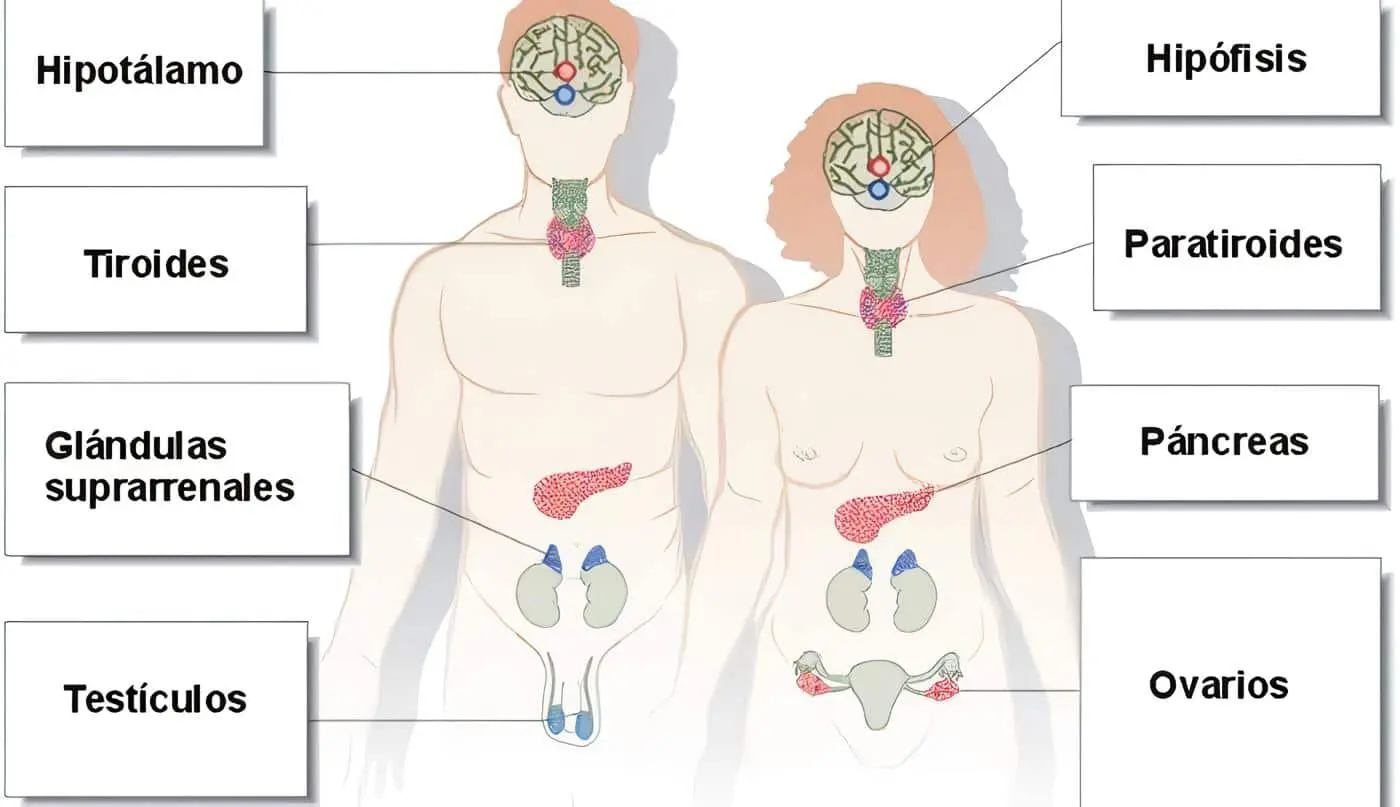
- 3) hacia el infundíbulo hipotalámico para controlar, al menos en parte, la mayoría de las funciones secretoras de la neurohipófisis y la adenohipófisis.
Los diversos mecanismos hipotalámicos encargados de controlar múltiples funciones del cuerpo:
- regular la presión arterial
- acción sobre la sed y la conservación del agua
- el apetito y el gasto de energía
- regulación de la temperatura
- control endocrino
Las áreas laterales resultan especialmente importantes para controlar la sed, el hambre y muchos de los impulsos emocionales.
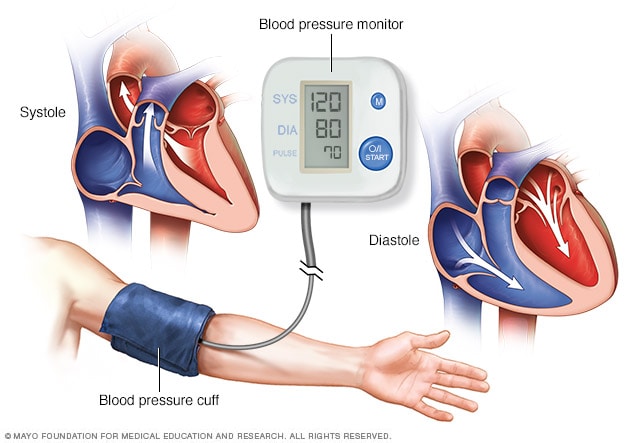

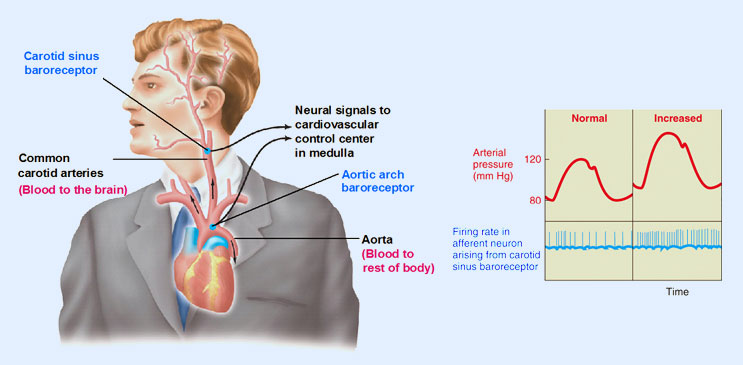
Regulación cardiovascular:
la estimulación del hipotálamo lateral y posterior eleva la presión arterial y la frecuencia cardíaca, mientras que la activación del área preóptica suele ejercer unos efectos opuestos, provocando una disminución de ambas variables.
Regulación de la temperatura corporal
El área preóptica, se ocupa de regular la temperatura corporal. Un incremento de esta variable en la sangre circulante a través de dicha área aumenta la actividad de las neuronas sensibles a la temperatura, mientras que su descenso la reduce.

Regulación del agua corporal
El hipotálamo regula el agua corporal por dos procedimientos:
- 1) originando la sensación de sed, lo que lleva a que el animal o la persona beban agua
- 2) controlando la excreción de agua en la orina. En el hipotálamo lateral está situada una zona denominada centro de la sed
- El control de la excreción renal de agua se encuentra asignado sobre todo a los núcleos supraópticos

Regulación digestiva y de la alimentación
Area hipotalámica lateral; la estimulación de diversas zonas hipotalámicas hace que un animal sienta un hambre enorme, un apetito voraz y un profundo deseo de buscar comida.
Las diversas zonas hipotalámicas controlan funciones vegetativas y endocrinas particulares. Las funciones de estas regiones aún no se entienden enteramente, de manera que las especificaciones facilitadas antes sobre las distintas áreas encargadas de las diferentes funciones hipotalámicas todavía son en parte provisionales.
Funciones de «recompensa» y de «castigo» cumplidas por el sistema límbico

Centros de recompensa
Por medio de esta técnica se ha descubierto que los principales centros de recompensa están situados a lo largo del trayecto del fascículo prosencefálico medial, sobre todo en los núcleos ventromedial y lateral del hipotálamo.

Centros de castigo
La estimulación de estas regiones hace que el animal muestre todos los signos de desagrado, miedo, terror, dolor, castigo y hasta enfermedad. Por medio de esta técnica se han descubierto las regiones más potentes encargadas de recibir el castigo y promover las tendencias de huida en la sustancia gris central del mesencéfalo que rodea al acueducto de Silvio y asciende por las zonas periventriculares del hipotálamo y el tálamo
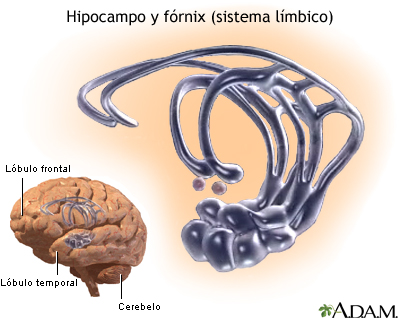
El hipocampo es la porción alargada de la corteza cerebral que se dobla hacia dentro para formar la cara ventral de gran parte del ventrículo lateral por su interior.
El hipocampo (y sus estructuras adyacentes de los lóbulos parietal y temporal, llamados en conjunto formación del hipocampo) posee numerosas conexiones con muchas porciones de la corteza cerebral, así como con las estructuras basales del sistema límbico (la amígdala, el hipotálamo, la región septal y los cuerpos mamilares), aunque sobre todo sean indirectas.
Otro rasgo propio del hipocampo es su posibilidad de volverse hiperexcitable.Probablemente una de las razones de esta hiperexcitabilidad de los hipocampos radique en que poseen un tipo de corteza diferente del que se encuentra en cualquier otro punto del telencéfalo.

El hipocampo surgió como una parte de la corteza olfatoria; una vez que estuviera asentada esta capacidad para tomar decisiones críticas, cabe pensar que el resto del encéfalo también comenzó a apelar al hipocampo con este fin. Por tanto, si su actividad indica que una información neuronal tiene importancia, es probable que su contenido resulte memorizado.
Sea cual sea el proceso que ocurre, sin el hipocampo la consolidación a largo plazo de los recuerdos de tipo verbal o de pensamiento simbólico es deficiente o no tiene lugar.