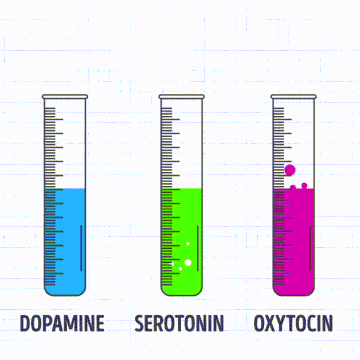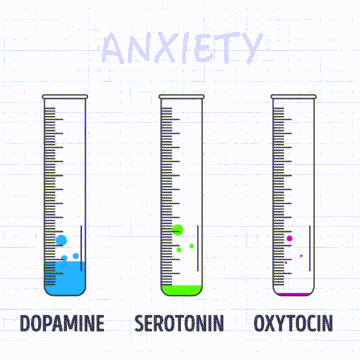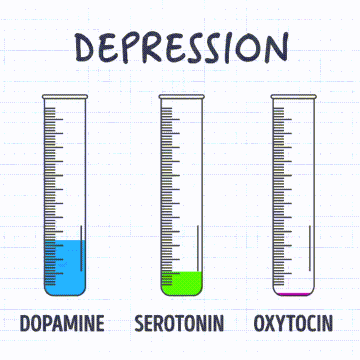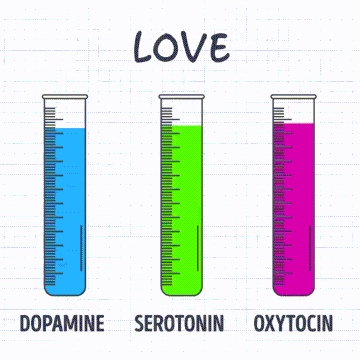
Receptores sensitivos, circuitos neuronales para el procesamiento de la información
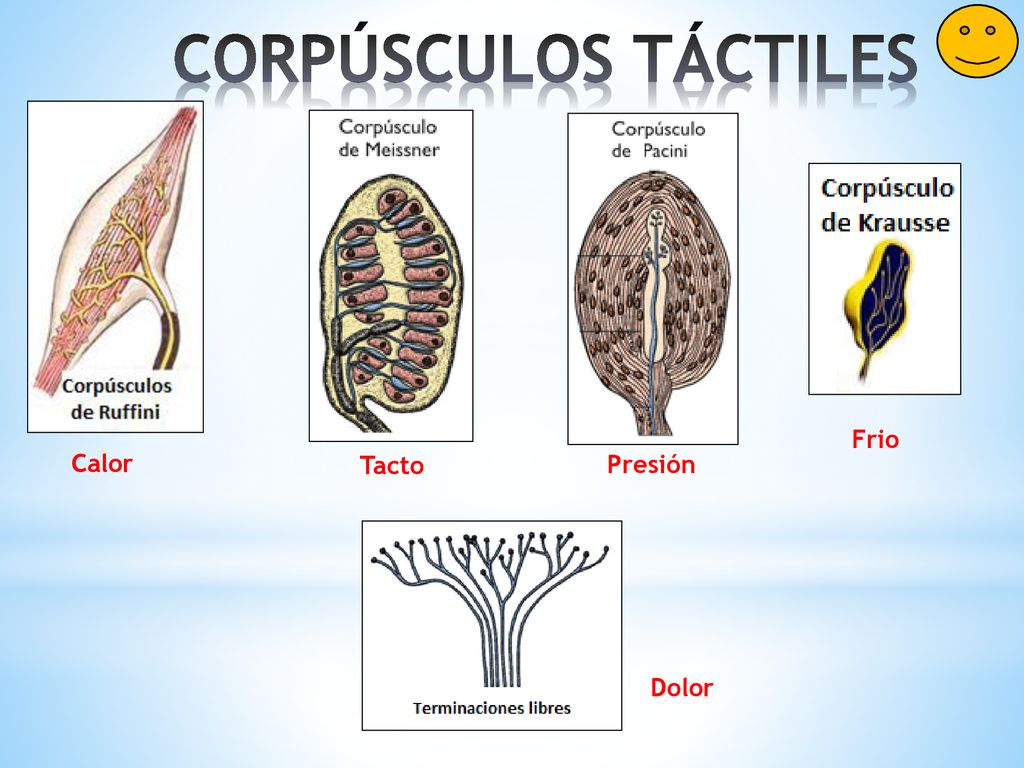
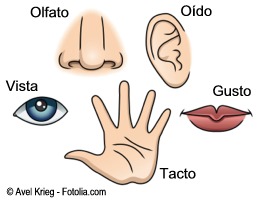
Tipos de receptores sensitivos y estímulos que detectan
tipos básicos de receptores sensitivos:
Mecanorreceptores
detectan la compresión mecánica o su estiramiento, o el de los tejidos adyacentes
TERMORECEPTORES
detectan los cambios en la temperatura
Nocirreceptores
detectan daños físicos o químicos que se producen en los tejidos
receptores electromagnéticos
detectan la luz en la retina ocular
quimiorreceptores
detectan el gusto en la boca, el olfato en la nariz, la cantidad de oxígeno en la sangre arterial, la osmolalidad de los líquidos corporales, la concentración de dióxido de carbono y otros factores que completen la bioquímica del organismo
La capacidad de los receptores sensitivos para detectar clases diferentes de estímulos sensitivos es debido a su SENSIBILIDAD DIFERENCIADA
Cada uno de los principales tipos sensitivos que podemos experimentar, dolor, tacto, visión, sonido, etc., se llama modalidad de sensación.
La especificidad de las fibras nerviosas para transmitir nada más que una modalidad de sensación se llama principio de la línea marcada.
Organización del sistema nervioso, funciones básicas de las sinapsis y neurotransmisores
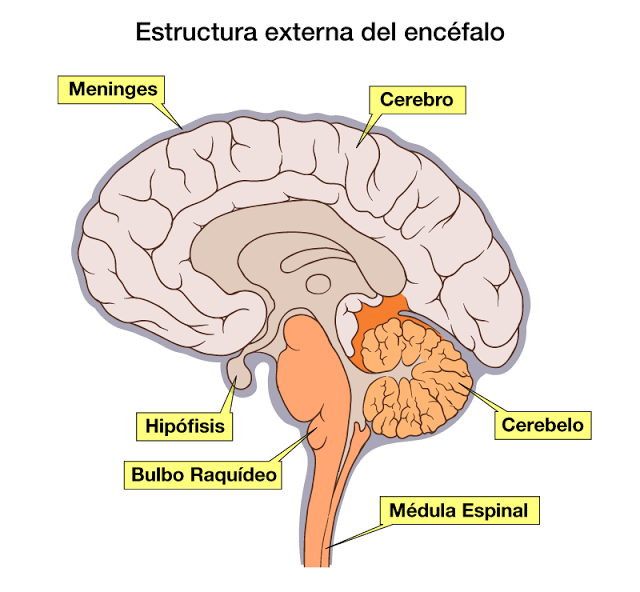
DISEÑO GENERAL DEL SISTEMA NERVIOSO
NEURONA:
Unidad funcional básica del sistema nervioso central:
posee la capacidad de recibir y decodificar información en forma de señales eléctricas y químicas, transmitiéndolas a otras células.
Las neuronas son las células más importantes, ya que son responsables de la transmisión de impulsos eléctricos a través del proceso de sinapsis, lo que constituye el principio del funcionamiento del cerebro.
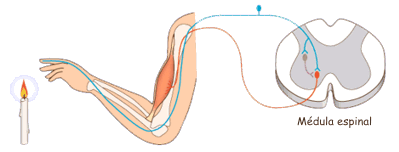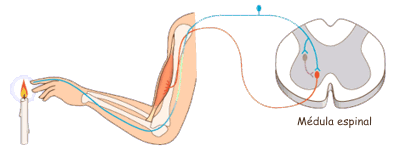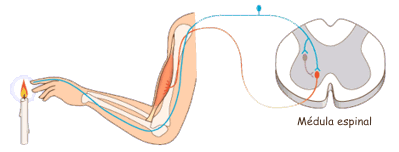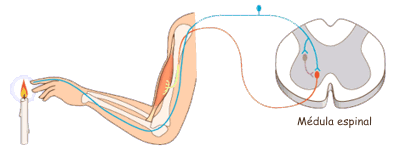
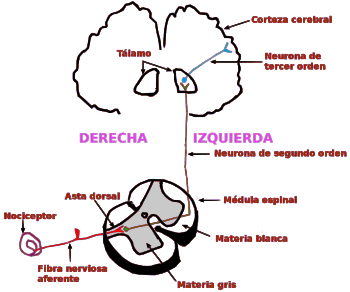
Las experiencias sensitivas excitan los receptores sensitivosy la informacion penetra en el sistema nervioso central a través de los nervios periféricos y se transporta de inmediato hasta múltiples zonas sensitivas en:
1) la médula espinal a todos sus niveles
2) la formación reticular del bulbo raquídeo, la protuberancia y el mesencéfalo en el encéfalo
sistema nervioso regula las diversas actividades del organismo.
1) la contracción de los músculos esqueléticos adecuados en todo el cuerpo
2) la contracción de la musculatura lisa de las vísceras, y
3) la secreción de sustancias químicas activas por parte de las glándulas exocrinas y endocrinas en muchas zonas del organismo
los musculos y las hormnoaas son llamados efectores ya que son las encargadas de efectuar las actividades llamadas funciones motoras del sistema nervioso:
eje nervioso motor «esquelético»= controlar la contracción de la musculatura esquelética.
sistema nervioso autónomo= encargado de controlar la musculatura lisa, las glándulas y otros sistemas corporales internos
Pueden controlarse a múltiples niveles del sistema nervioso central, como por ejemplo:
1) la médula espinal
2) la formación reticular del bulbo raquídeo, la protuberancia y el mesencéfalo
se le denomina función integradora del sistema nervioso a la llegada de información sensitiva importante excitante a la mente que de inmediato resulta encauzada hacia las regiones motoras e integradoras oportunas del encéfalo para suscitar las respuestas deseadas.
La memoria es una función del cerebro que permite al organismo codificar, almacenar y recuperar la información del pasado. Algunas teorías afirman que surge como resultado de las conexiones sinápticas repetitivas entre las neuronas, lo que crea redes neuronales.
Cada vez que determinados tipos de señales sensitivas atraviesan una secuencia de sinapsis, estas adquieren una mayor capacidad para transmitir ese mismo tipo de señal la próxima vez, situación que llamamos facilitación. Después de que las señales sensitivas hayan recorrido las sinapsis en multitud de ocasiones, su facilitación es tan profunda que las señales generadas dentro del propio encéfalo también pueden originar la transmisión de impulsos a lo largo de la misma serie de sinapsis, incluso cuando no haya sido estimulada su entrada sensitiva. Este proceso otorga a la persona una percepción de estar experimentando sensaciones originales, aunque únicamente se trate de recuerdos.
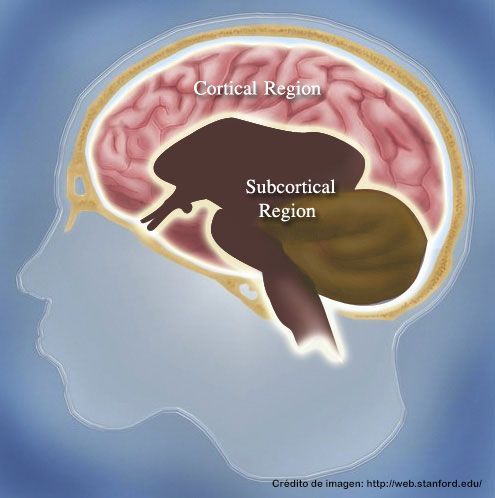
Principales niveles de función del sistema nervioso central
los principales niveles del sistema nervioso central que presentan unas características funcionales específicas son tres:
1) el nivel medular
2) el nivel encefálico inferior o subcortical
3) el nivel encefálico superior o cortical.
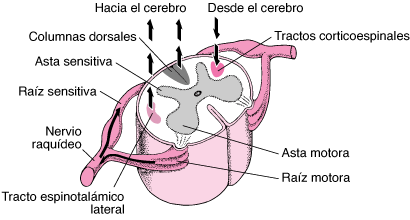
Nivel medular
los circuitos neuronales de la médula pueden originar:
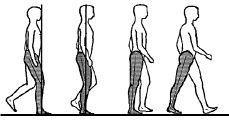
1) los movimientos de la marcha
2) reflejos para retirar una parte del organismo de los objetos dolorosos
3) reflejos para poner rígidas las piernas para sostener el tronco en contra de la gravedad
4) reflejos que controlan los vasos sanguíneos locales, los movimientos digestivos o la excreción urinaria.
Es el nivel que destapa todo un mundo de información almacenada para que la mente la use.
Sin su concurso, el funcionamiento de los centros encefálicos inferiores a menudo es impreciso. El inmenso depósito de información cortical suele convertir estas funciones en operaciones determinativas y precisas.
La información recorre el sistema nervioso central sobre todo bajo la forma de potenciales de acción nerviosos, llamados simplemente impulsos nerviosos.
La sinapsis química es el sitio en que células vecinas se comunican entre sí a través de mensajes químicos, los neurotransmisores. A pesar del enorme número de sinapsis químicas que existen en el sistema nervioso y de la amplia variedad estructural que ellas ofrecen, en la organización de este tipo de sinapsis se pueden reconocer los mismos elementos básicos. Hay un elemento presináptico representado por un terminal nervioso, o una varicosidad o por el polo de liberación de mensajes químicos, que se observa en algunos tipos celulares, como algunas células sensoriales.
Hasta hoy se han descubierto más de 40 neurotransmisores importantes.
Entre las mejor conocidas figuran las siguientes:
acetilcolina
noradrenalina
adrenalina
histamina
ácido γ-aminobutírico (GABA)
glicina,
serotonina
glutamato.
Las sinapsis químicas poseen una característica sumamente importante que las convierte en un elemento muy conveniente para transmitir la mayor parte de las señales en el sistema nervioso. Esta característica hace posible que siempre conduzcan las señales en un solo sentido: es decir, desde la neurona que segrega el neurotransmisor, denominada neurona presináptica, hasta la neurona sobre la que actúa el transmisor, llamada neurona postsináptica. Este fenómeno es el principio de la conducción unidireccional de las sinapsis químicas y se aleja bastante de la conducción a través de las sinapsis eléctricas, que muchas veces transmiten señales en ambos sentidos.
axón, que se extiende desde el soma hacia un nervio periférico para abandonar la médula espinal
las dendritas, que constituyen una gran cantidad de prolongaciones ramificadas
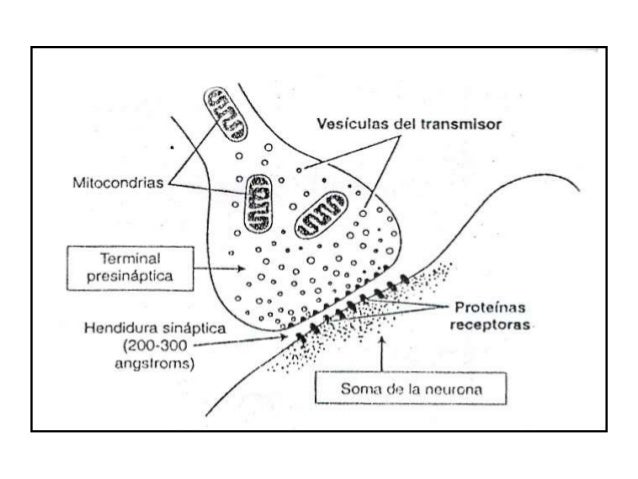
El terminal está separado del soma neuronal postsináptico por una hendidura sináptica en él existen dos estructuras internas de importancia para la función excitadora o inhibidora de la sinapsis:
las vesículas transmisoras y las mitocondrias.
Las vesículas transmisoras contienen el neurotransmisor que, cuando se libera a la hendidura sináptica, excita o inhibe la neurona postsináptica. Excita la neurona postsináptica si la membrana neuronal posee receptores excitadores e inhibe la neurona si contiene receptores inhibidores.
Las mitocondrias aportan trifosfato de adenosina (ATP), que a su vez suministra energía para sintetizar más sustancia transmisora.
Cuando un potencial de acción la despolariza, estos canales se abren y permiten la entrada en el terminal de un número importante de iones calcio.
los iones calcio llegan al terminal presináptico, se unen a unas moléculas proteicas especiales situadas sobre la cara interna de la membrana presináptica, llamadas puntos de liberación. A su vez, este enlace suscita la apertura de los puntos de liberación a través de la membrana, y así permite que unas pocas vesículas transmisoras suelten su contenido hacia la hendidura después de cada potencial de acción.
función de las «proteínas receptoras:
La membrana de la neurona postsináptica contiene una gran cantidad de proteínas receptora
La activación de los receptores controla la apertura de los canales iónicos en la célula postsináptica según una de las dos formas siguientes:
1) por activación de los canales iónicos para permitir el paso de determinados tipos de iones a través de la membrana
2) mediante la activación de un «segundo mensajero» que en vez de un canal iónico es una molécula que protruye hacia el citoplasma celular y activa una sustancia o más en el seno de la neurona postsináptica.
receptores ionotrópicos= receptores de neurotransmisores que activan directamente los canales iónicos
receptores metabotrópicos.= receptores de neurotransmisores que actúan a través de sistemas de segundos mensajeros
Sistema de «segundo mensajero» en la neurona postsináptica:
Los procesos de memoria, requieren la producción de unos cambios prolongados en las neuronas durante segundos y hasta meses después de la desaparición de la sustancia transmisora inicial. Los canales iónicos no son idóneos para originar una variación prolongada en las neuronas postsinápticas, porque se cierran en cuestión de milisegundos una vez desaparece la sustancia transmisora.
Transmisores de acción rápida y molécula pequeña/ neuropeptidos
Los transmisores de acción rápida y molécula pequeña son los que producen las respuestas más inmediatas del sistema nervioso
se sintetizan en el citoplasma del terminal presináptico y las numerosas vesículas transmisoras presentes a este nivel los absorben por transporte activo, se liberan en la hendidura sináptica sobre los receptores de la membrana en la neurona postsináptica incrementando o disminuiyendo la conductancia que presentan los canales iónicos
tienen acciones lentas formandose en los ribosomas del soma neuronal, una característica importante de los neuropéptidos es que a menudo ocasionan acciones mucho más duraderas. Algunas de ellas consisten en el cierre prolongado de los canales de calcio, los cambios persistentes en la maquinaria metabólica de las células, en la activación o la desactivación de genes específicos dentro del núcleo celular, o en alteraciones a largo plazo de la cantidad de receptores excitadores o inhibidores.
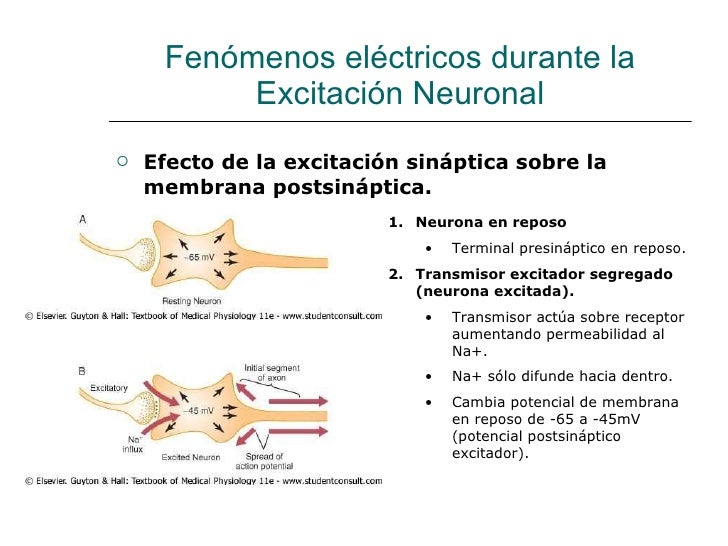
Fenómenos eléctricos durante la excitación neuronal
El descenso del voltaje hasta un nivel menos negativo vuelve más excitable la membrana de la neurona, mientras que su aumento hasta un nivel más negativo la hace menos excitable. Este mecanismo es el fundamento de los dos modos de acción en la neurona
El interior del soma neuronal contiene una solución electrolítica muy conductora, el líquido intracelular de la neurona Por tanto, todo cambio en el potencial de cualquier zona del líquido dentro del soma suscita un cambio casi exactamente igual en el potencial de los demás puntos de su interior.
Generación de potenciales de acción en el segmento inicial del axón a su salida de la neurona: umbral de excitación:
La razón principal para que este sea el punto de origen del potencial de acción reside en que el soma posee en su membrana relativamente pocos canales de sodio dependientes de voltaje, lo que complica la apertura por parte del PPSE del número necesario para desencadenar un potencial de acción.
Una vez que comienza el potencial de acción, viaja en sentido periférico a lo largo del axón y normalmente también en sentido retrógrado hacia el soma.
Fenómenos eléctricos durante la inhibición neuronal
Las sinapsis inhibidoras sobre todo abren canales de cloruro, lo que permite el paso sin problemas de estos iones.
La apertura de los canales de potasio dejará que estos iones de carga positiva se desplacen hacia el exterior y esto también volverá más negativo de lo normal el potencial de membrana interno. Así pues, la entrada de cloruro más la salida de potasio elevan el grado de negatividad intracelular, lo que se denomina hiperpolarización
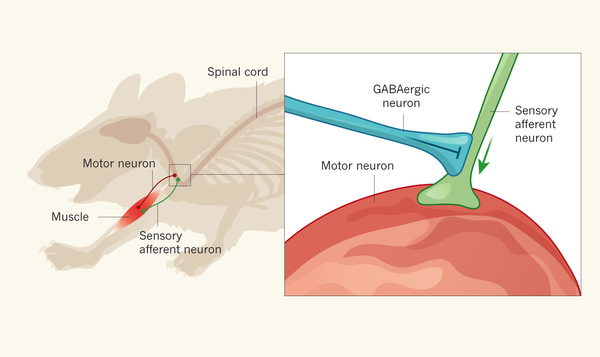
La inhibición presináptica está ocasionada por la liberación de una sustancia inhibidora en las inmediaciones de las fibrillas nerviosas presinápticas antes de que sus propias terminaciones acaben sobre la neurona postsináptica. En la mayoría de los casos, la sustancia transmisora inhibidora es GABA (ácido γ-aminobutírico).Ocurre en muchas de las vías sensitivas del sistema nervioso.
«Sumación espacial» en las neuronas: umbral de disparo
Este fenómeno reside en que la cantidad de sustancia transmisora liberada por un terminal aislado para originar un PPSE normalmente no supera los 0,5 a 1 mV, en vez de los 10 a 20 mV necesarios en general para alcanzar el umbral de excitación.
Cada vez que dispara un terminal presináptico, la sustancia transmisora liberada abre los canales de membrana a lo sumo durante 1 ms más o menos. Una segunda apertura de estos mismos elementos puede incrementar el potencial postsináptico hasta un nivel aún mayor y cuanto más alta sea la velocidad de estimulación, mayor se volverá el potencial postsináptico.
Funciones especiales de las dendritas para excitar a las neuronas
La mayoría de las dendritas no llegan a transmitir potenciales de acción debido a que sus membranas poseen relativamente pocos canales de sodio dependientes de voltaje, y sus umbrales de excitación son demasiado elevados para producir potenciales de acción.
Este proceso significa la propagación directa de una corriente eléctrica por conducción de iones en los líquidos de las dendritas pero sin la generación de potenciales de acción.
Relación del estado de excitación de la neurona con la frecuencia de descarga
Si en un momento determinado el grado de excitación es más alto que el de inhibición, entonces se dice que existe un estado excitador. A la inversa, si es mayor la inhibición que la excitación, lo que se dice es que hay un estado inhibidor
Cuando las sinapsis excitadoras reciben estímulos repetidos a un ritmo elevado, el número de descargas de la neurona postsináptica es muy alto al principio, pero la frecuencia de disparo va bajando progresivamente en los milisegundos o segundos sucesivos
Se sabe que muchos fármacos aumentan la excitabilidad de las neuronas y otros la disminuyen. Por ejemplo, la cafeína, la teofilina y la teobromina, que están presentes en el café, el té y el chocolate, respectivamente, incrementan la excitabilidad neuronal, se supone que al rebajar el umbral de excitación en las células.
Durante la transmisión de una señal neuronal desde una neurona presináptica hasta otra postsináptica, se consume cierta cantidad de tiempo en el proceso siguiente:
1) emisión de la sustancia transmisora por el terminal presináptico;
2) difusión del transmisor hacia la membrana neuronal postsináptica;
3) acción del transmisor sobre el receptor de la membrana;
4) intervención del receptor para aumentar la permeabilidad de la membrana
5) entrada del sodio por difusión para elevar el PPSE hasta un nivel suficientemente alto como para desencadenar un potencial de acción.
Formado por fibras mucho menores, habitualmente de 1 a 5 μm de diámetro y de solo 20 a 500 μm de longitud.
Tipos de músculo liso:
1) dimensiones físicas
2) organización en fascículos o láminas
3) respuesta a diferentes tipos de estímulos
4) características de la inervación
5) función.
En aras de la simplicidad, el músculo liso en general se puede dividir:
músculo liso multiunitario
Formado por fibras musculares lisas separadas y discretas. Cada una de las fibras actúa independientemente de las demás y con frecuencia está inervada por una única terminación nerviosa, como ocurre en las fibras musculares esqueléticas.
Cada una de las fibras se puede contraer independientemente de las demás, y que su control se ejerce principalmente por señales nerviosas.
músculo liso unitario (o monounitario).
se denomina músculo liso sincitial o músculo liso visceral.
. Este tipo de músculo liso también se conoce como músculo liso sincitial debido a sus interconexiones sincitiales entre las fibras. También se denomina músculo liso visceral porque se encuentra en la pared de la mayor parte de las vísceras del cuerpo, por ejemplo:
el aparato digestivo
las vías biliares
los uréteres,
el útero y
muchos vasos sanguíneos.
Mecanismo contráctil en el músculo liso
Base química de la contracción del músculo liso :}
El músculo liso contiene filamentos tanto de actina como de miosina, que tienen características químicas similares a los filamentos de actina y miosina del músculo esquelético. No contiene el complejo de troponina necesario para el control de la contracción del músculo esquelético, de modo que el mecanismo de control de la contracción es diferente.
Base física de la contracción del músculo liso
La fuerza de contracción se transmite de unas células a otras principalmente a través de estos enlaces.
Esta configuración permite que la miosina tire de un filamento de actina en una dirección en un lado a la vez que tira de otro filamento de actina en la dirección opuesta en el otro lado. La utilidad de esta organización es que permite que las células musculares lisas se contraigan hasta el 80% de su longitud, en lugar de estar limitadas a menos del 30%, como ocurre en el músculo esquelético.
Comparación de la contracción del músculo liso con la contracción del músculo estriado
MUSCULO LISO
contracciones del músculo liso son contracciones tónicas prolongadas, que a veces duran horas o incluso días.
es mucho más lenta que en el músculo esquelético
Baja necesidad de energía para mantener la contracción del músculo liso
Lentitud del inicio de la contracción y relajación del tejido muscular liso total
La fuerza máxima de contracción muscular es a menudo mayor en el músculo liso
El mecanismo de «cerrojo» facilita el mantenimiento prolongado de las contracciones del músculo liso
su capacidad de recuperar casi su fuerza de contracción original segundos a minutos después de que haya sido alargado o acortado.
llamado así por presentar estriaciones
la mayor parte de él se asocia al esqueleto,
funciona bajo control voluntario ya que se encuentra inervado por el sistema nervioso somático.
Está constituido por largas células multinucleadas, cuyos núcleos se localizan en la periferia. Este tipo muscular se encuentra en la lengua, la faringe, en el segmento superior del esófago y en la porción lumbar del diafragma, además de los músculos extrínsecos del ojo y en toda la musculatura de las extremidades y del tronco
Aproximadamente el 40% del cuerpo es músculo esquelético, y tal vez otro 10% es músculo liso y cardíaco
Anatomía fisiológica del músculo esquelético :
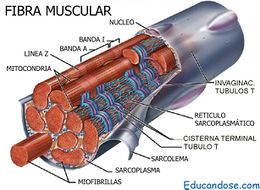
Fibras del músculo esquelético
Todos los músculos esqueléticos están formados por numerosas fibras cuyo diámetro varía entre 10 y 80 μm. Cada una de estas fibras está formada por subunidades cada vez más pequeñas
Todas las fibras, excepto alrededor de un 2%, habitualmente están inervadas por una sola terminación nerviosa localizada cerca del punto medio de la fibra
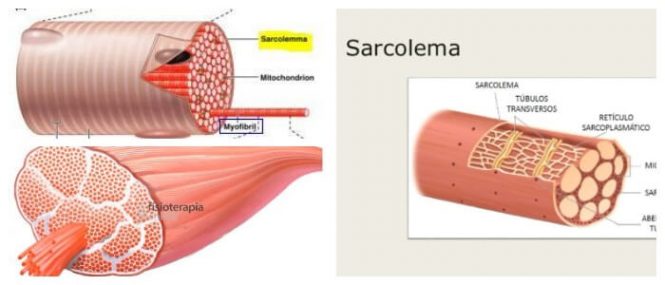
El sarcolema:
es una fina membrana que envuelve a una fibra musculoesquelética denominada membrana plasmática, y una cubierta externa formada por una capa delgada de material polisacárido que contiene numerosas fibrillas delgadas de colágeno.
Las miofibrillas
Cada fibra muscular contiene varios cientos a varios miles de miofibrillas, que se representan en la vista en sección transversal. Cada miofibrilla está formada por aproximadamente 1.500 filamentos de miosina y 3.000 filamentos de actina adyacentes entre sí, que son grandes moléculas proteicas polimerizadas responsables de la contracción muscular real
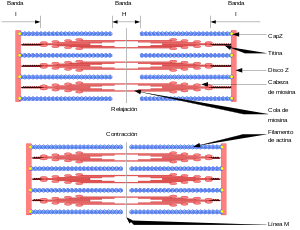
Las bandas claras contienen solo filamentos de actina y se denominan bandas I porque son isótropas a la luz polarizada. Las bandas oscuras contienen filamentos de miosina, así como los extremos de los filamentos de actina en el punto en el que se superponen con la miosina, y se denominan bandas A porque son anisótropas a la luz polarizada.
El disco Z, que está formado por proteínas filamentosas distintas de los filamentos de actina y miosina, atraviesa las miofibrillas y también pasa desde unas miofibrillas a otras, uniéndolas entre sí a lo largo de toda la longitud de la fibra muscular.
Las moléculas filamentosas de titina mantienen en su lugar los filamentos de miosina y actina:
La relación de yuxtaposición entre los filamentos de miosina y de actina se mantiene por medio de un gran número de moléculas filamentosas de una proteína denominada titina
Estas moléculas elásticas de titina actúan como armazón que mantiene en su posición los filamentos de miosina y de actina, de modo que funcione la maquinaria contráctil del sarcómero.
. La molécula de titina también parece actuar como molde para la formación inicial de porciones de los filamentos contráctiles del sarcómero, especialmente los filamentos de miosina.
El sarcoplasma
Las muchas miofibrillas de cada fibra muscular están yuxtapuestas suspendidas en la fibra muscular. Los espacios entre las miofibrillas están llenos de un líquido intracelular denominado sarcoplasma, que contiene grandes cantidades de potasio, magnesio y fosfato, además de múltiples enzimas proteicas.
El retículo sarcoplásmico:
En el sarcoplasma que rodea a las miofibrillas de todas las fibras musculares también hay un extenso retículo tiene una organización especial que es muy importante para regular el almacenamiento, la liberación y la recaptación de calcio y, por tanto, para controlar la contracción muscular.
Mecanismo general de la contracción muscular
El inicio y la ejecución de la contracción muscular se producen en las siguientes etapas secuenciales:
1. Un potencial de acción viaja a lo largo de una fibra motora hasta sus terminales sobre las fibras musculares.
2. En cada terminal, el nervio secreta una pequeña cantidad de la sustancia neurotransmisora acetilcolina.
3. La acetilcolina actúa en una zona local de la membrana de la fibra muscular para abrir múltiples canales de cationes «activados por acetilcolina» a través de moléculas proteicas que flotan en la membrana.
4. La apertura de los canales activados por acetilcolina permite que grandes cantidades de iones sodio difundan hacia el interior de la membrana de la fibra muscular. Esta acción provoca una despolarización local que, a su vez, conduce a la apertura de los canales de sodio activados por el voltaje, que inicia un potencial de acción en la membrana.
5. El potencial de acción viaja a lo largo de la membrana de la fibra muscular de la misma manera que los potenciales de acción viajan a lo largo de las membranas de las fibras nerviosas.
6. El potencial de acción despolariza la membrana muscular, y buena parte de la electricidad del potencial de acción fluye a través del centro de la fibra muscular, donde hace que el retículo sarcoplásmico libere grandes cantidades de iones calcio que se han almacenado en el interior de este retículo.
7. Los iones calcio inician fuerzas de atracción entre los filamentos de actina y miosina, haciendo que se deslicen unos sobre otros en sentido longitudinal, lo que constituye el proceso contráctil.
8. Después de una fracción de segundo los iones calcio son bombeados de nuevo hacia el retículo sarcoplásmico por una bomba de Ca++ de la membrana y permanecen almacenados en el retículo hasta que llega un nuevo potencial de acción muscular; esta retirada de los iones calcio desde las miofibrillas hace que cese la contracción muscular.
Mecanismo molecular de la contracción muscular
La contracción muscular se produce por un mecanismo de deslizamiento de los filamentos.
En el estado relajado, los extremos de los filamentos de actina que se extienden entre dos discos Z sucesivos apenas comienzan a superponerse entre sí.
En el estado contraído estos filamentos de actina han sido traccionados hacia dentro entre los filamentos de miosina, de modo que sus extremos se superponen entre sí en su máxima extensión
Esta acción está producida por las fuerzas que se generan por la interacción de los puentes cruzados que van desde los filamentos de miosina a los filamentos de actina
Características moleculares de los filamentos contráctiles
Los filamentos de miosina están compuestos por múltiples moléculas de miosina.
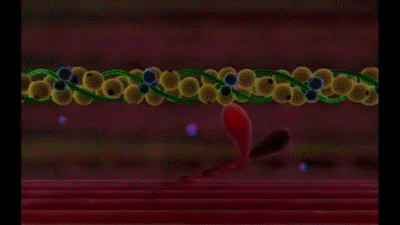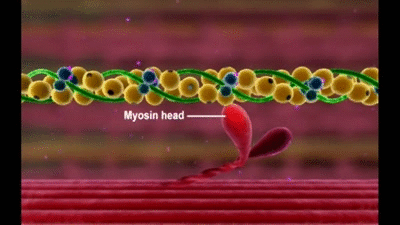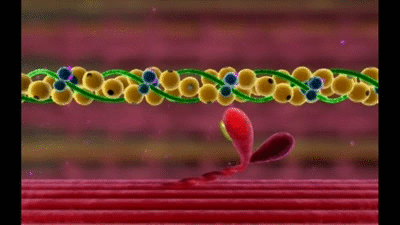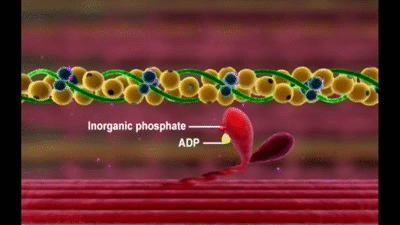
La molécula de miosina está formada por seis cadenas polipeptídicas, dos cadenas pesadas, cada una de las cuales tiene un peso molecular de aproximadamente 200.000, y cuatro cadenas ligeras, que tienen un peso molecular de aproximadamente 20.000 cada una. Las dos cadenas pesadas se enrollan entre sí en espiral para formar una hélice doble, que se denomina cola de la molécula de miosina. Un extremo de cada una de estas cadenas se pliega bilateralmente para formar una estructura polipeptídica globular denominada cabeza de la miosina. Así, hay dos cabezas libres en un extremo de la molécula de miosina de doble hélice. Las cuatro cadenas ligeras también forman parte de la cabeza de la miosina, dos en cada cabeza. Estas cadenas ligeras ayudan a controlar la función de la cabeza durante la contracción muscular.
Cada puente cruzado es flexible en dos puntos denominados bisagras, una en el punto en el que el brazo sale del cuerpo del filamento de miosina y la otra en el punto en el que la cabeza se une al brazo.
Actividad adenosina trifosfatasa de la cabeza de miosina :
Otra característica de la cabeza de la miosina que es esencial para la contracción muscular es que actúa como una enzima adenosina trifosfatasa (ATPasa)
Esta propiedad permite que la cabeza escinda el ATP y que utilice la energía procedente del enlace fosfato de alta energía del ATP para aportar energía al proceso de la contracción.
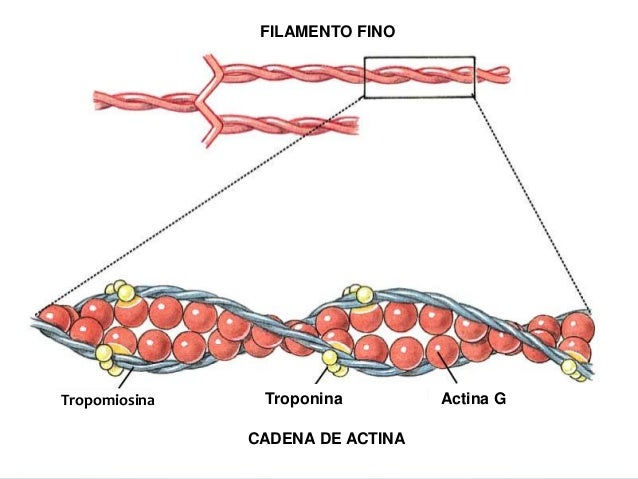
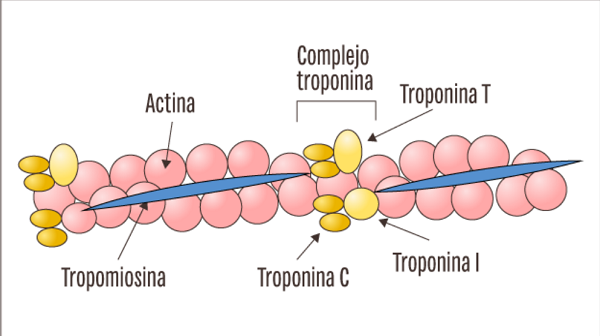
Los filamentos de actina están formados por actina, tropomiosina y troponina:
El esqueleto del filamento de actina es una molécula de la proteína F-actina bicatenaria, Las dos hebras están enroscadas en una hélice de la misma manera que la molécula de miosina.
tropomiosina
El filamento de actina también contiene otra proteína, la tropomiosina. Cada molécula de tropomiosina tiene un peso molecular de 70.000 y una longitud de 40 nm. Estas moléculas están enrolladas en espiral alrededor de los lados de la hélice de F-actina. En estado de reposo las moléculas de tropomiosina recubren los puntos activos de las hebras de actina, de modo que no se puede producir atracción entre los filamentos de actina y de miosina para producir la contracción.
Troponina
Estas moléculas proteicas son en realidad complejos de tres subunidades proteicas unidas entre sí de manera laxa, cada una de las cuales tiene una función específica en el control de la contracción muscular. Una de las subunidades (troponina I) tiene una gran afinidad por la actina, otra (troponina T) por la tropomiosina y la tercera (troponina C) por los iones calcio.
Hay potenciales eléctricos a través de las membranas de prácticamente todas las células del cuerpo.
En otros tipos de células, como las células glandulares, los macrófagos y las células ciliadas, los cambios locales de los potenciales de membrana también activan muchas de las funciones de las células
La ecuación de Nernst describe la relación del potencial de difusión con la diferencia de concentración de iones a través de una membrana:
El nivel del potencial de difusión a través de una membrana que se opone exactamente a la difusión neta de un ion particular a través de la membrana se denomina potencial de Nernst para ese ion.
Cuando se utiliza esta fórmula habitualmente se asume que el potencial del líquido extracelular que está fuera de la membrana se mantiene a un nivel de potencial cero, y que el potencial de Nernst es el potencial que está en el interior de la membrana.
La ecuación de Goldman se utiliza para calcular el potencial de difusión cuando la membrana es permeable a varios iones diferentes:
Cuando una membrana es permeable a varios iones diferentes, el potencial de difusión que se genera depende de tres factores:
1) la polaridad de la carga eléctrica de cada uno de los iones
2) la permeabilidad de la membrana (P) a cada uno de los iones
3) las concentraciones (C) de los respectivos iones en el interior (i) y en el exterior (e) de la membrana
Da el potencial de membrana calculado en el interior de la membrana cuando participan dos iones positivos univalentes, sodio (Na+) y potasio (K+), y un ion negativo univalente, cloruro (Cl–).
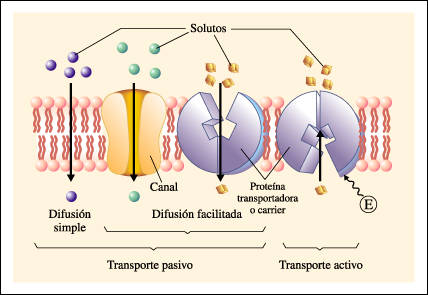
TRANSPORTE DE SUSTANCIAS A TRAVÉS DE LAS MEMBRANAS CELULARES
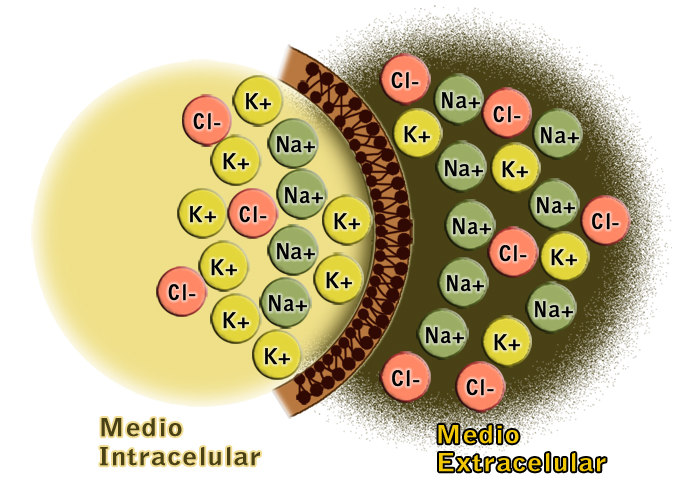
Las diferencias entre el liquido intracelular y extracelular que son muy importantes para la vida de la célula son:
Liquido extracelular
Contiene una gran cantidad de sodio
Contiene una pequeña cantidad de potasio
contiene una gran cantidad de iones cloruro
Liquido intracelular
contiene una gran cantidad de potasio
contiene una pequeña cantidad de sodio
contiene muy poca cantidad de iones cloruro
contiene una gran concentración de fosfatos y proteínas
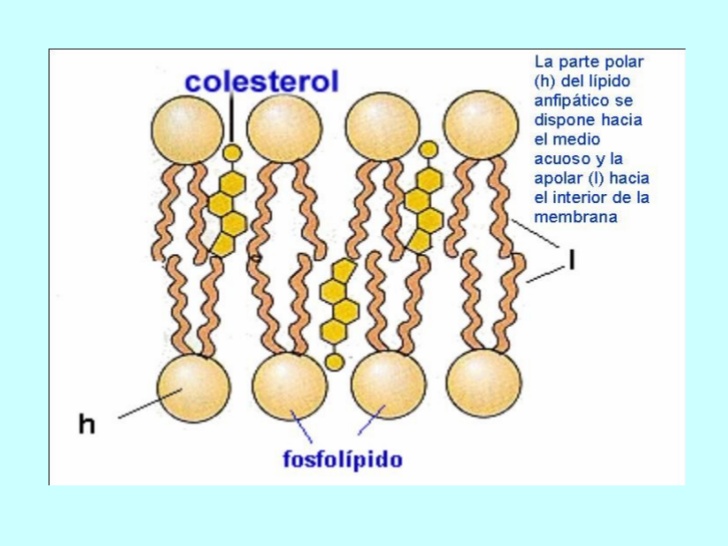
LA MEMBRANA CELULAR CONSISTE EN UNA BICAPA LIPÍDICA CON PROTEÍNAS DE TRANSPORTE DE LA MEMBRANA CELULAR
La bicapa lipídica no es miscible con el líquido extracelular ni con el líquido intracelular. Por tanto, constituye una barrera frente al movimiento de moléculas de agua y de sustancias insolubles entre los compartimientos del líquido extracelular e intracelular.
Las moléculas proteicas de la membrana tienen unas propiedades totalmente diferentes para transportar sustancias.
PROTEÍNAS TRANSPORTADORAS: Son proteínas las cuales su estructura interrumpen la continuidad de la bicapa lipidica y constituyen una ruta alterna a través de la membrana
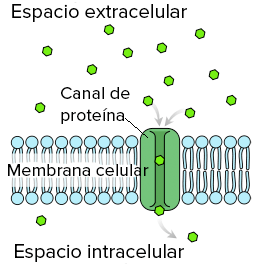
PROTEÍNAS DE LOS CANALES: Proteínas que contienen espacios acuosos en todo el trayecto del interior de la molécula, permitiendo el paso libre de agua, iones y moléculas seleccionadas
PROTEÍNAS TRANSPORTADORAS: Proteínas que se unen a las moléculas que se van a transportar la cual sufre cambios conformacionales, realizando el desplazo de las sustancias a través de intersticios de las proteinas hasta el otro lado
Procesos básicos en los que se produce el transporte de la membrana celular ya sea directamente a través de la bicapa lipídica o a través de las proteínas
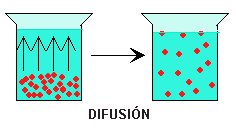
la difusión se refiere a un movimiento molecular aleatorio de las sustancias molécula a molécula, a través de espacios intermoleculares de la membrana o en combinación con una proteína transportadora. La energía necesaria para que se produzca la difusión es la energía del movimiento cinético normal de la materia.
el transporte activo se refiere al movimiento de iones o de otras sustancias a través de la membrana en combinación con una proteína transportadora de tal manera que la proteína transportadora hace que la sustancia se mueva contra un gradiente de energía
Difusión
Debido a que todas las moléculas iones y sustancias disueltas están en movimiento constante, de modo que cada partícula se mueve de manera completamente independiente a este movimiento continuo de moléculas entre sí en los líquidos o los gases.
La difusión a través de la membrana celular se divide en dos subtipos:
difusión simple
difusión facilitada.
Movimiento cinético de las moléculas o de los iones se produce a través de una abertura de la membrana o a través de espacios intermoleculares sin ninguna interacción con las proteínas transportadoras de la membrana.
DIFUSIÓN DE SUSTANCIAS LIPOSOLUBLES A TRAVÉS DE LA BICAPA LIPÍDICA:
Liposolubulidad es unfactor importante que determina la rapidez con la que una sistancia se difunde a traves de la bicapa lipidica
aquellas sustancias que pueden disolverse directamente en la bicapa lipidica son
oxigeno
anhídrido carbónico
alcoholes
nitrógeno
Difusión de agua y de otras moléculas insolubles en lípidos a través de canales proteicos:
debido a que el agua es insoluble en los lipidos, debe pasar rapidamente por los canales de las moleculas proteicas que pentetran en todo el espesor de la memebrana.
Necesitan unas en específicos llamadas «acuaporinas» que permiten el paso rápido de agua a través de la membrana, existen al menos 13 tipos diferentes.
Al igual que otras moléculas son insolubles en lipidos pueden utilizar los canales de los poros proteicos .
Los poros están compuestos por proteínas de membranas celulares integrales que forman tubos abiertos a través de la membrana y que están siempre abiertos; las caracteristicas de un poro que le permite su selectividad dandole paso a ciertas moleculas son :
Diámetro del poro
Cargas eléctricas
su forma
Enlaces químicos situados a lo largo de las superficies internas
Estos canales se distinguen por:
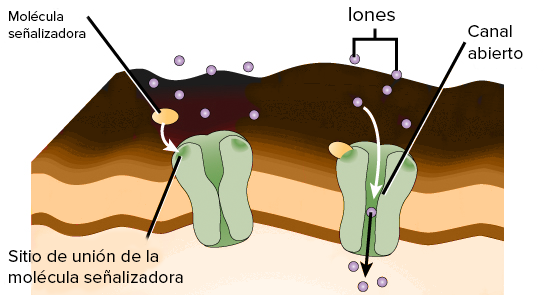
Son permeables de manera selectiva a ciertas sustancias
los canales se pueden abrir o cerrar por compuertas que son reguladas por señales eléctricas (canales activados por el voltaje) o sustancias químicas que se unen a las proteínas de canales (canales activados por ligandos).
Permeabilidad selectiva de los canales proteicos
un ejemplo de esto seria:
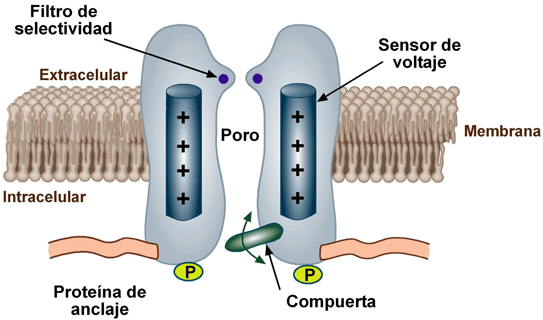
Se descubrió que los canales de potasio tienen una estructura tetramérica consistente en cuatro subunidades proteicas idénticas que rodean a un poro central (fig. 4-4). En la parte superior del poro del canal se distribuyen bucles de poro que forman un estrecho filtro de selectividad. Como revestimiento del filtro de selectividad hay oxígenos de carbonilo. Cuando los iones potasio hidratados entran en el filtro de selectividad, interaccionan con los oxígenos de carbonilo y envuelven la mayoría de sus moléculas de agua ligadas, lo que permite que los iones potasio deshidratados pasen a través del canal. Sin embargo, los oxígenos de carbonilo están demasiado separados para permitir su interacción estrecha con los iones sodio, más pequeños, que de este modo son excluidos en la práctica por el filtro de selectividad y no pueden pasar a través del poro.
La existencia diferentes filtros de selectividad determinan, la especificidad de los diversos canales para cationes o aniones o para iones determinados como:
sodio (Na+)
potasio (K+)
calcio (Ca++),
Activación de los canales proteicos
El proceso de activacion de canales proporciona un medio para controlar la permeabilidad iónica de los canales.
La apertura y el cierre de las compuertas están controlados de dos maneras principales:
ACTIVACION POR VOLTAJE
La conformación molecular de la compuerta o de sus enlaces químicos responde al potencial eléctrico que se establece a través de la membrana celular.
ACTIVACION POR LIGANDOS
Se abren por la unión de una sustancia química (un ligando) a la proteína, que produce un cambio conformacional o un cambio de los enlaces químicos de la molécula de la proteína que abre o cierra la compuerta.
Estado abierto frente a estado cerrado de los canales activados:
Se debe tomar en cuenta que en el canal se conduce la corriente según un mecanismo de «todo o nada». Es decir, la compuerta del canal se abre súbitamente y después se cierra súbitamente.
A un potencial de voltaje dado, el canal puede permanecer cerrado todo o casi todo el tiempo.
Otro nivel de voltaje puede permanecer abierto todo o la mayor parte del tiempo.
Voltajes intermedios las compuertas tienden a abrirse y cerrarse súbitamente de manera intermitente, lo que da un flujo medio de corriente que está en algún punto entre el mínimo y el máximo.
Método del pinzamiento zonal de membrana (patch-clamp) para el registro del flujo de las corrientes iónicas a través de canales aislados:
El método del «pinzamiento zonal de membrana» (patch-clamp) para el registro del flujo de una corriente iónica a través de canales proteicos aislados.
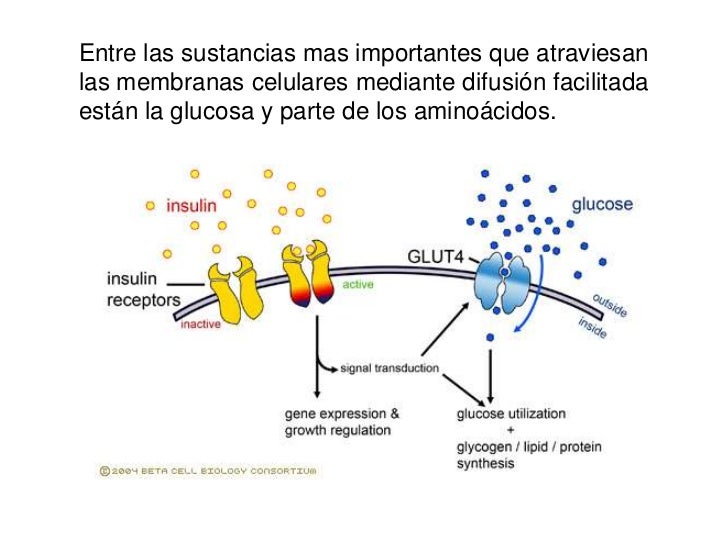
También se denomina difusión mediada por un transportador porque una sustancia que se transporta a través de la membrana con la ayuda de una proteína transportadora específica para contribuir al transporte.
Hay una característica que hace que la difusión facilitada difiera de la difusión simple:
a medida que aumenta la concentración de la sustancia que difunde, la velocidad de la difusión simple sigue aumentando de manera proporcional, aunque en el caso de la difusión facilitada la velocidad de la difusión no puede aumentar por encima del nivel de la Vmáx.
sustancias que atraviesan las membranas celulares mediante difusión facilitada están la glucosa y la mayoría de los aminoácidos.
El encargado de transportar la glucosa se le conoce como GLUT en este caso de la se han descubierto en varios tejidos al menos 14 miembros de una familia de proteínas de membrana. También pueden transportar otros monosacáridos que tienen estructuras similares a la glucosa como:
Importante es la velocidad neta de difusión de una sustancia en la dirección deseada. Esta velocidad neta está determinada por varios factores.
Hacia adentro:
La velocidad a la que la sustancia difunde hacia dentro es proporcional a la concentración de las moléculas en el exterior, porque esta concentración determina cuántas moléculas chocan contra el exterior de la membrana cada segundo.
Hacia afuera:
la velocidad a la que las moléculas difunden hacia afuera es proporcional a su concentración en el interior de la membrana.
Efecto del potencial eléctrico de membrana sobre la difusión de iones: el «potencial de Nernst»
Cuando la diferencia de concentración se hace lo suficientemente elevada, los dos efectos se contrarrestan entre sí. A la temperatura corporal normal (37 °C), la diferencia eléctrica que permitirá que se alcance el equilibrio entre una diferencia de concentración dada de iones univalentes, como los iones Na+, se puede determinar a partir de la fórmula siguiente, que se denomina ecuación de Nernst:
El agua al ser la sustancia más abundante que se difunde a través de la membrana celular se difunde en ambas direcciones a través de la membrana . Estas cantidades que se difunde en ambas direcciones está equilibrada de manera tan precisa que se produce un movimiento neto cero de agua, manteniéndose el volumen celular constante.
El proceso de movimiento neto del agua que se debe a la producción de una diferencia de la concentración del agua se denomina ósmosis
La cantidad de presión necesaria para detener la ósmosis se denomina presión osmótica
Importancia del número de partículas osmóticas (concentración molar) en la determinación de la presión osmótica
La presión osmótica que ejercen las partículas de una solución se determina por el número de partículas por unidad de volumen del líquido
La razón es porque todas las partículas de una solución , ejercen, la misma cantidad de presión contra la membrana.
El factor que determina la presión osmótica de una solución es la concentración de la solución en función del número de partículas
«Osmolalidad»: el osmol
Para expresar la concentración de una solución en función del número de partículas se utiliza la unidad denominada osmol en lugar de los gramos.
Un osmol es el peso molecular-gramo de un soluto osmóticamente activo.
Así, se dice que una solución que tiene 1 osmol de soluto disuelto por cada kilogramo de agua tiene una osmolalidad de 1 osmol por kilogramo
La osmolaridad normal de los líquidos extracelular e intracelular es de aproximadamente 300 mosmol por kilogramo de agua.
Relación entre osmolalidad y presión osmótica:
A la temperatura corporal normal, 37 °C, una concentración de un osmol por litro producirá una presión osmótica de 19.300 mmHg en la solución.
La concentración de 1 mosmol por litro es equivalente a una presión osmótica de 19,3 mmHg.
La multiplicación de este valor por la concentración 300 miliosmolar de los líquidos corporales da una presión osmótica calculada total de los líquidos corporales de 5.790 mmHg.
El término «osmolaridad»
es la concentración osmolar expresada en osmoles por litro de solución
más práctico medir la osmolaridad, por lo tanto esta es la práctica habitual en casi todos los estudios fisiológicos.
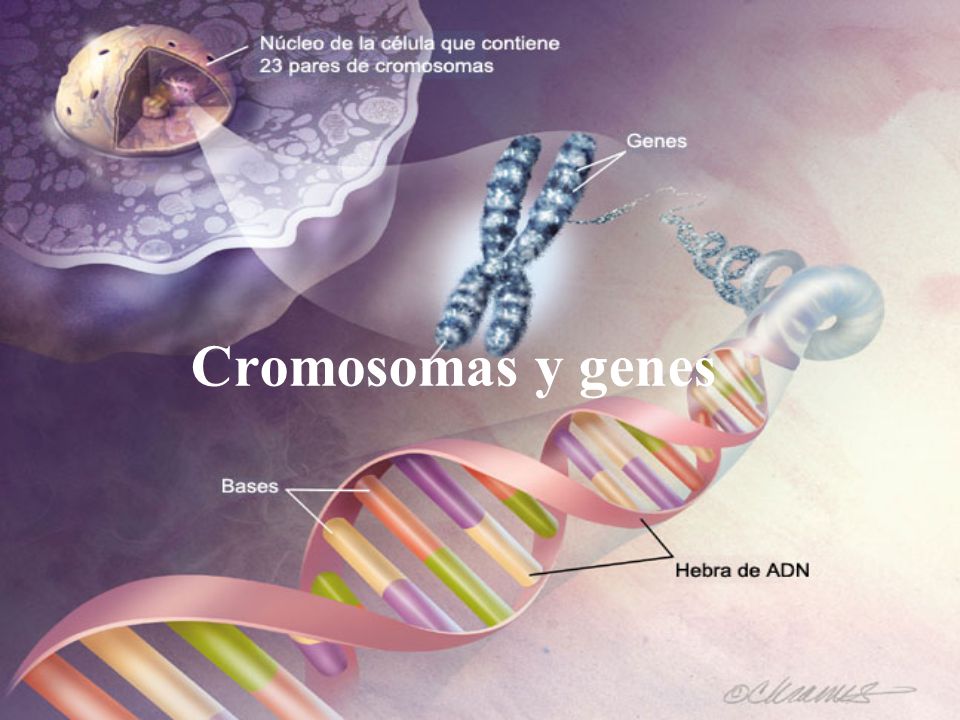
Control genético de la síntesis proteica, las funciones de la célula y la reproducción celular
Los genes que se situan en el nucleo de las celulas pueden:
Controlar la herencia de padres a hijos.
Controlan la función cotidiana de las células del organismo.
Controlan las funciones de la célula determinando que sustancias se sintetizaran.
Cada gen esta compuesto por:
Acido desoxirribonucleico que controla la formación de otro acido nucleico (ARN)
Dispersa por toda la célula para controlar la formacion de una proteina especifica
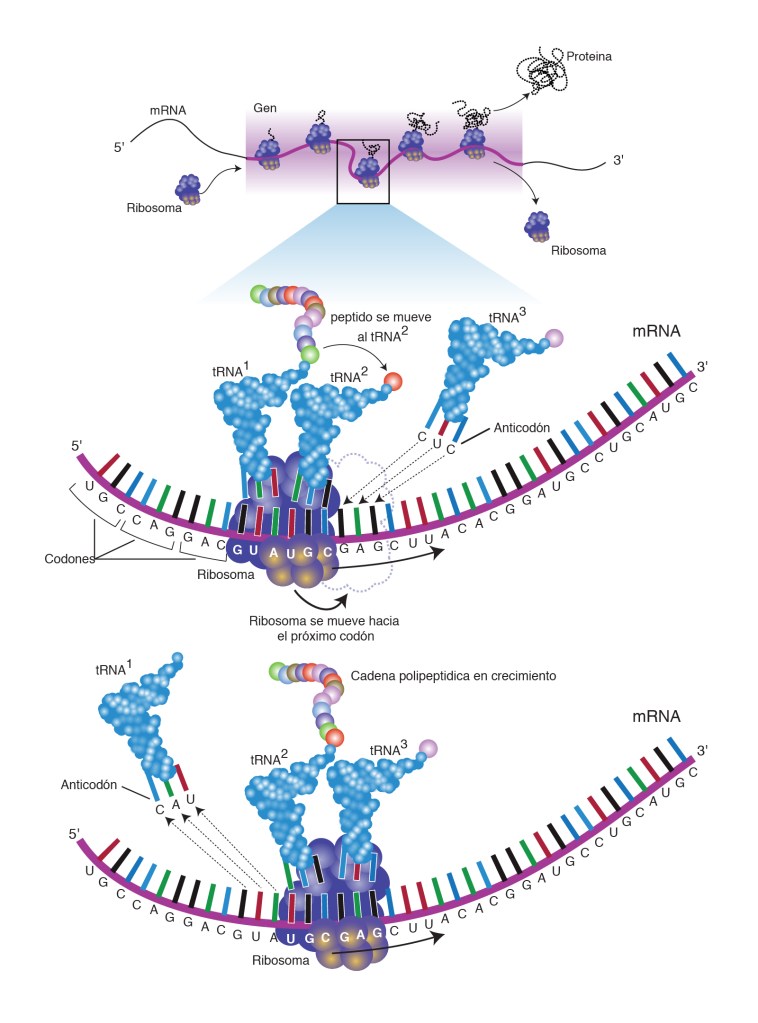
Expresión Génica: Se le llama así al proceso completo desde la transcripción del código genético en el núcleo hasta la traducción del código del ARN para la formación de proteínas en el citoplasma celular.
Se estima una cantidad de 30,000 genes diferentes en cada celula , con la capacidad de formar un gran numero de proteinas celulares diferentes.
Algunas de las cuales son proteínas estructurales que asociado a varios lipidos e hidratos de carbono, forman las estructuras de los distintos orgánulos intracelulares o mas bien proteínas tipo Enzimas que catalizan distintas reacciones oxidativas que aportan energía a la célula y la favorecen en la síntesis de los productos químicos de la célula como lipidos, glucogeno y ATP.
Proteína estructural
Enzimas
Los genes en el núcleo celular controlan la síntesis de las proteínas.
Los genes se encuentran en el núcleo unido por sus extremos, formando moléculas de doble hélice largas de ADN,
Los compuestos químicos básicos que forman el ADN incluyen:
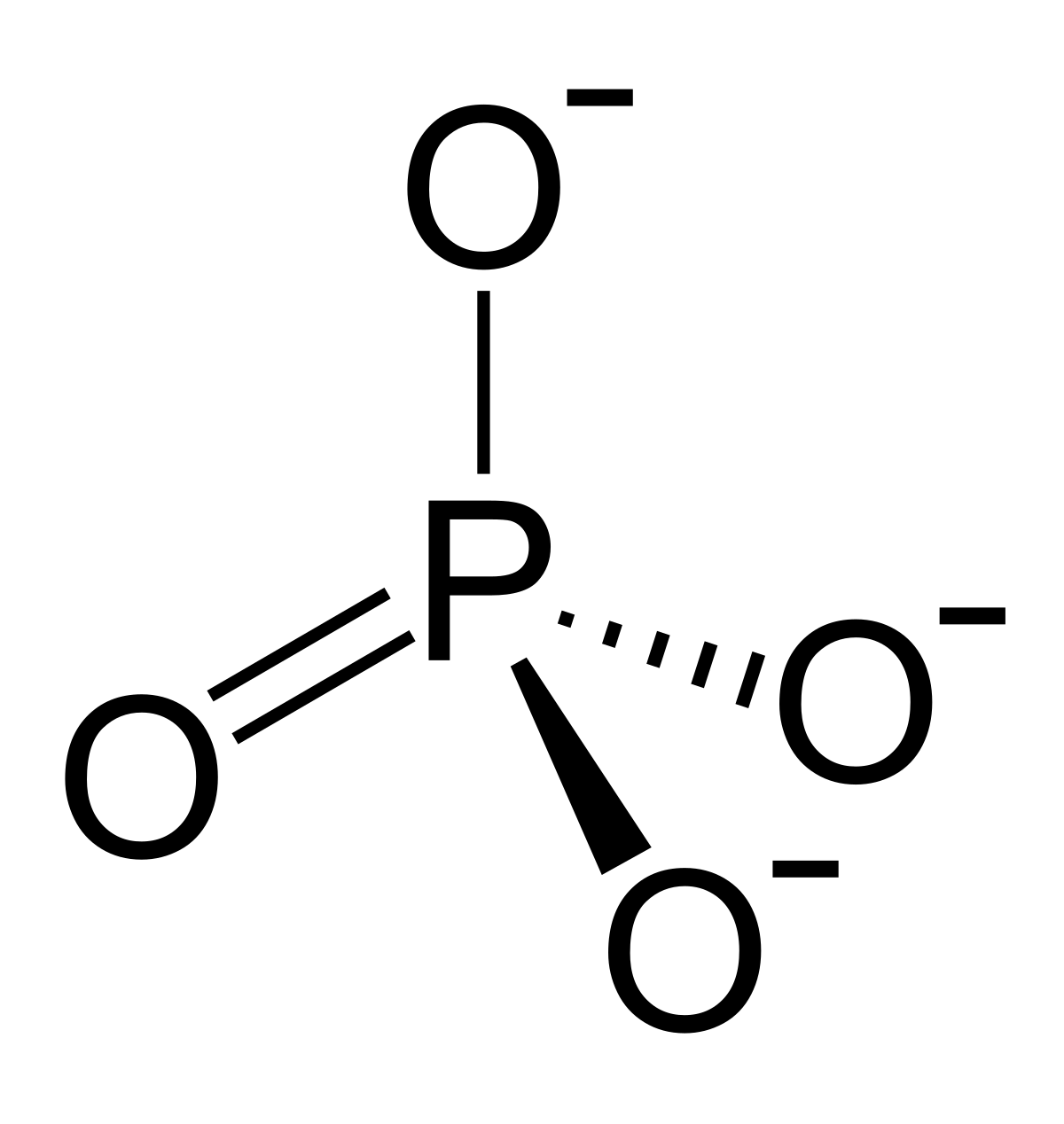
ÁCIDO FOSFÓRICO:
AZÚCAR PENTOSA:
BASE NITROGENADA:
El ácido fosforico y la desoxirribosa forman las hebras helicoidales que sirven de soporte para la molécula de ADN, mientras que las bases nitrogenadas se apoyan entre las dos hebras y se conectan entre si.
Para la formación del ADN el primer paso es combinar una molécula de ácido fosfórico, una molécula de desoxirribosa y una de las cuatro bases para formar un nucleotido ácido. De esta forma se pueden formar 4 nucleótidos distintos.
Un gran numero de nucleotidos se unen ente si para formar las dos hebras del ADN y estas dos hebras se unen laxamente mediante enlaces débiles;en esta organización se deben tomar ciertos aspectos como:
El esqueleto de ADN esta formado por la alternación de desoxirribosa y el ácido fosfórico.
Las bases purina y pirimidina se unen a los lados de las moléculas de desoxirribosa.
Las hebras respectivas de ADN se mantiene unidas mediante enlaces de hidrógeno.
Cada base purínica de Adenina de una hebra siempre se une con una base Pirimidínica de Timina de la otra
Cada base purínica de Guanina siempre se une con una base pirimidínica de Citosina.
En cada vuelta completa de la hélice de la molécula de ADN hay 10 pares de nucleótidos
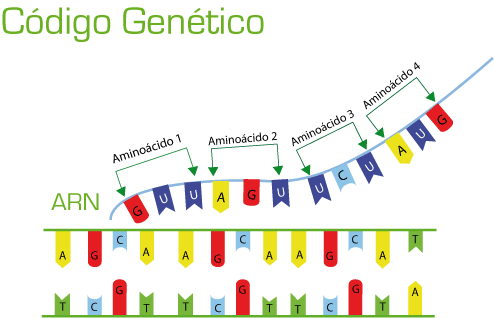
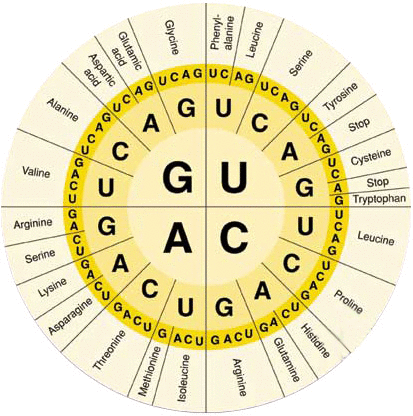
La capacidad para controlar la formación de proteínas en la célula es una de las importancias del ADN, esto se consigue mediante un código genético.
Se le conoce como código genético al escindir las hebras de la molécula de ADN exponiendo las bases purinicas y pirimidinicas proyectándose a cada lado de la hebra .
El código genético consta de «tripletes»sucesivos de bases que componen una palabra del código, estos controlan la secuencia de aminoácidos en una molécula proteica que la célula debe sintetizar.
Los tripletes son responsables de la colocación sucesiva de los aminoácidos en una molécula de proteína de nueva formación.
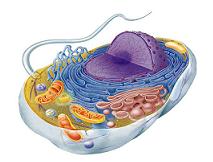
Cada célula del ser humano es una estructura viva que puede sobrevivir durante meses o incluso muchos años, siempre que los líquidos de su entorno contengan los nutrientes apropiados. Aportan la estructura de los tejidos y los órganos del cuerpo, ingieren los nutrientes y los convierten en energía, al igual contienen el código hereditario del organismo que controla las sustancias sintetizadas por las células y les permite realizar copias de si mismas.
Organización de la célula
Las dos partes mas importantes de una célula son el núcleo y el citoplasma, separadas entre si por una membrana nuclear mientras que el encargado de separar el citoplasma de los líquidos circundantes se le llama membrana plasmática o membrana celular.
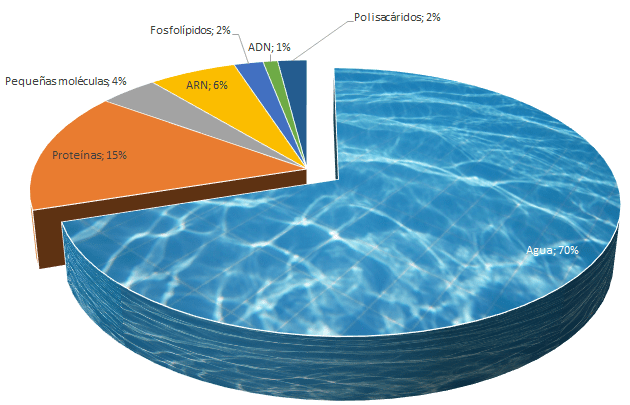
Todas las sustancias y elementos que componen a la célula en conjunto reciben el nombre de protoplasma. Compuesto principalmente por:
Agua
Electrolitos
Proteínas
Lipidos
Hidratos de carbono
AGUA: Principal medio liquido de la célula, presente en la mayoría de las células excepto los adipocitos ; en este se encuentran disueltos muchos de los componentes químicos y las reacciones químicas tienen lugar entre los productos químicos disueltos o en la superficie de las membranas
Iones: Son los productos químicos inorgánicos de las reacciones celulares , necesarios para el funcionamiento de algunos de los mecanismos de control celular .
Potasio
Magnesio
Fosfato
Sulfato
Bicarbonato
y en cantidades mas pequeñas:
Sodio
Cloruro
Calcio
Proteínas: Son las sustancias mas abundantes de las células, constituyendo entre el 10% y 20%, se distinguen entre:
Proteína estructural
Proteína funcional
Proteína estructural: Presente en las células en forma de filamentos largos que son polímeros de muchos moléculas proteicas individuales. Algunos de sus usos importantes son:
(filamentos intracelulares)Formación de microtubulos que proporcionan los «citoesqueletos» de organelos celulares como los cilios,axones nerviosos y husos mitoticos
(comportimiento extracelular)Las proteínas fibrilares se encuentran en las fibras de colágeno y elastina del tejido conjuntivo y en las paredes de los vasos sanguíneos,tendones, ligamentos.
Proteína funcional: compuestas por combinaciones de pocas moléculas en forma tubular-globular. Son principalmente las enzimas móviles dentro del liquido celular y muchas de ellas se encuentran adheridas a la estructura membranosa realizando la catálisis de las reacciones químicas intracelulares especificas.
Lipidos: son varios tipos de sustancias que se agrupan por ser en común solubles en disolventes grasos, los lípidos importantes son :
Fosfolípidos
Colesterol
Juntos suponiendo un 2% de la masa total de la celular, debido a que son insolubles al agua son usados para formar las barreras de la membrana celular y de la membrana intracelular que separan los compartimientos celulares.
trigliceridos
Los triglicéridos componen el 95% de los adipocitos, la grasa almacenada en estas células representa el principal almacén del organismo de nutrientes energéticos que después se pueden disolver y usarse para proporcionar energía siempre que el organismo la necesite.
Hidratos de carbono:Forman parte de las moléculas glucoproteicas y tienen una importante función en la nutrición de la célula; la media de las células humanas que presentan una reserva de hidratos de carbono son :
1% de la masa total de una célula
3% en las células musculares
6% en células hepáticas
En el liquido extracelular los hidratos de carbono se encuentran en forma de glucosa de manera accesible a la célula, mientras que en la parte intraceluar se almacena una pequeña cantidad de hidratos de carbono en forma de glucógeno (polimero insoluble de glucosa que se puede despolimerizar rápidamente para aportar a la célula la energía que necesiten»)
Esta es la primera entrada de mi nuevo blog. Acabo de empezar a escribir este nuevo blog. ¡Mantente al día para leer más entradas! Suscríbete a continuación para recibir notificaciones cuando publique nuevo contenido.
Esto es una entrada de muestra, originalmente publicada como parte de Blogging University. Regístrate en uno de nuestros diez programas y empieza tu blog con buen pie.
Hoy vas a publicar una entrada. No te preocupes por el aspecto de tu blog. Tampoco te preocupes si todavía no le has puesto un nombre o si todo esto te agobia un poco. Tan solo haz clic en el botón «Nueva entrada» y explícanos por qué estás aquí.
¿Por qué es necesario?
Porque proporciona contexto a los nuevos lectores. ¿A qué te dedicas? ¿Por qué deberían leer tu blog?
Porque te ayudará a concentrarte en tus propias ideas sobre tu blog y en lo que quieres hacer con él.
La entrada puede ser corta o larga, una introducción de tu vida o una declaración de los objetivos del blog, un manifiesto de cara al futuro o una breve descripción del tipo de cosas que pretendes publicar.
Te ofrecemos algunos consejos útiles para ayudarte a empezar:
¿Por qué publicas tus entradas en lugar de escribir en un diario personal?
¿Sobre qué temas crees que escribirás?
¿Con quién te gustaría conectar a través de tu blog?
Si tu blog resulta ser un éxito a lo largo del próximo año, ¿qué objetivo te gustaría lograr?
No tienes por qué atarte a las decisiones que tomes ahora. Lo bueno de los blogs es que evolucionan constantemente a medida que aprendemos nuevas cosas, crecemos e interactuamos los unos con los otros. Pero está bien saber dónde y por qué empezaste, y plasmar tus objetivos puede darte más ideas para las entradas que quieres publicar.
¿No sabes por dónde empezar? Tan solo escribe lo primero que se te ocurra. Anne Lamott, autora de un libro sobre cómo escribir que nos encanta, afirma que debemos permitirnos escribir un «primer borrador de mierda». Anne está en lo cierto: tan solo tienes que empezar a escribir, y ya te encargarás de editarlo más tarde.
Cuando todo esté listo para publicarse, asigna entre tres y cinco etiquetas a la entrada que describan el centro de atención de tu blog: escritura, fotografía, ficción, educación, comida, coches, películas, deportes… ¡Lo que sea! Estas etiquetas ayudarán a los usuarios interesados en tus temas a encontrarte en el Lector. Una de las etiquetas debe ser «zerotohero», para que los nuevos blogueros también puedan encontrarte.




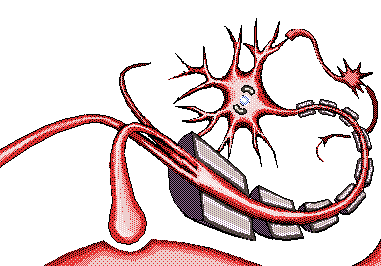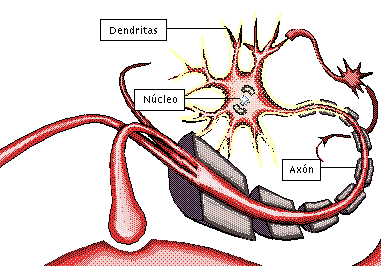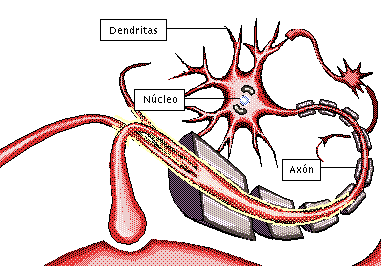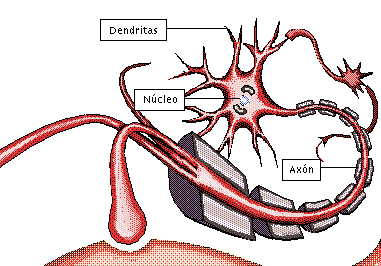
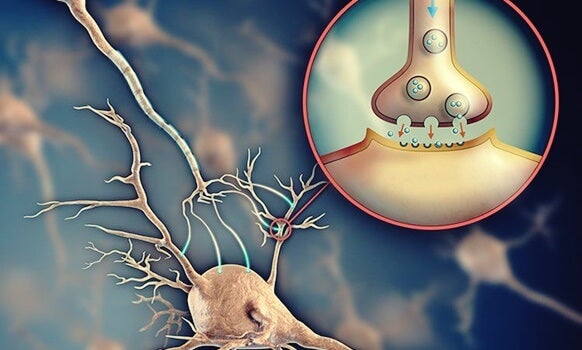 Las neuronas son las células más importantes, ya que son responsables de la transmisión de impulsos eléctricos a través del proceso de sinapsis, lo que constituye el principio del funcionamiento del cerebro.
Las neuronas son las células más importantes, ya que son responsables de la transmisión de impulsos eléctricos a través del proceso de sinapsis, lo que constituye el principio del funcionamiento del cerebro.